|
| Appunti universita |
|
|
| Appunti universita |
|
| Visite: 2116 | Gradito: |
Leggi anche appunti:VibrioniVIBRIONI ¨ Bacilli ¨ GRAM Patologia da agenti fisiciPATOLOGIA DA AGENTI FISICI USTIONI L' ustione è il processo patologico Dermatiti bolloseDERMATITI BOLLOSE Si tratta di affezioni caratterizzate clinicamente da bolle |
 |
 |
Infiammazione delle vie aeree
L'infiammazione cronica delle vie aeree è una caratteristica delle malattie respiratorie di tipo ostruttivo. L'infiammazione interessa tutte le vie aeree, ma in particolar modo i bronchi di medie dimensioni. Diversi elementi cellulari e numerosi mediatori risultano avere un ruolo nella modulazione della risposta infiammatoria nelle vie respiratorie (Wenzel, 2003).
Cellule infiammatorie
Eosinofili. Gli eosinofili possiedono un ampio spettro di proprietà biologiche, come la capacità di rilasciare proteine tossiche dai loro granuli, radicali liberi dell'ossigeno, eicosanoidi (leucotrieni solfo-peptidici) (Busse at al., 1994), fattore attivante le piastrine (PAF), citochine analoghe a quelle prodotte dai Th2 (Ying et al., 1995; Broide et al., 1992) e una varietà di fattori di crescita (Weller, 1991; Venge et al., 1987). La loro capacità di secernere mediatori può essere attivata da meccanismi sia immunologici che non immunologici (Venge et al., 1987). Gli eosinofili attivati possono far iniziare la contrazione del muscolo liscio nelle vie aeree umane (Weller, 1991), aumentare la permeabilità microvascolare (Collins et al., 1993) e indurre iperresponsività bronchiale (Leff, 1994). Nei campioni bioptici bronchiali, prelevati da soggetti con asma cronico, è stato rilevato un aumentato numero di eosinofili attivati, prevalentemente al di sotto della membrana basale. La maggior parte delle persone con asma allergico o non allergico, comprese quelle con asma lieve, hanno eosinofili nel lume e nella parete delle vie aeree.
Mastociti. I mastociti si localizzano nei bronchi sia di soggetti normali sia di quelli affetti da patologie respiratorie Oltre a rilasciare mediatori autacoidi, i mastociti sono un'importante fonte di citochine e di proteasi neutre, specialmente di triptasi, che agiscono su substrati proteici, come ad esempio i recettori attivati da proteasi (Pesci et al., 1993; Koshino et al., 1995). I mastociti attivati rilasciano mediatori broncocostrittori (istamina, cistenil-leucotrieni, prostaglandina D2) (Galli et al., 2005). Queste cellule sono attivate da allergeni, tramite i recettori IgE ad alta affinità, o da stimoli osmotici (broncocostrizione indotta da esercizio fisico). L'aumentato numero di mastociti nella muscolatura liscia delle vie aeree è stato messo in relazione all'aumentata iperreattività bronchiale (Robinson et al.,
Linfociti I linfociti T, presenti in numero elevato nelle vie aeree, rilasciano citochine specifiche, comprese IL-4, IL-5, IL-9 e IL-13, che orchestrano l'infiammazione eosinofilica e la produzione di IgE dai linfociti B (Larche et al., 2003). Un aumento nell'attività delle cellule Th2 può essere dovuto in parte ad una riduzione delle cellule T regolatrici che normalmente inibiscono le cellule Th2. Ci può anche essere un aumento delle cellule iNKT, che rilasciano grandi quantità di citochine Th1 e Th2 (Akbari et al., 2006).
Le cellule dendritiche, catturano gli allergeni dalla superficie delle vie aeree e migrano ai linfonodi regionali, dove interagiscono con le cellule T regolatrici ed infine stimolano la produzione delle cellule Th2 da cellule T naïve98.
Neutrofili. I neutrofili sono aumentati nelle vie aeree e nell'espettorato di pazienti con asma grave e negli asmatici fumatori, ma il ruolo fisiopatologico di queste cellule è incerto ed il loro aumento può persino essere dovuto alla terapia con corticosteroidi (Ying et al., 1995). I neutrofili polimorfonucleati sono stati considerati per lungo tempo come cellule in stadio di differenziazione terminale, incapaci di sintesi proteica e deputate solo a ruolo di effettori passivi dell'infiammazione attraverso la fagocitosi e il rilascio di enzimi preformati e di composti citotossici (Leckie et al., 2000; Lloyd et al.,1992; Wenzel et al., 1997). Tuttavia i neutrofili possono sintetizzare un'ampia varietà di enzimi tra cui proteasi, che degradano la matrice extracellulare (per esempio la MMP-9 e l'elastasi), specie reattive dell'ossigeno, citochine e chemochine come IL-1β, TNF-α, IL-6 e IL-8.
Macrofagi I macrofagi sono aumentati nelle vie aeree del paziente affetto da asma e BPCO e possono essere attivati da allergeni tramite i recettori a bassa- affinità delle IgE, per liberare i mediatori infiammatori e le citochine che amplificano la risposta infiammatoria (Peters-Golden, 2004). I macrofagi tissutali hanno la capacità di secernere un'ampia varietà di prodotti, molti dei quali svolgono un ruolo di rilievo nei processi di danno e riparazione (Nathan, 1987). Essi sintetizzano e secernono l'attivatore del plasminogeno e un gruppo di metalloproteasi che possono degradare varie macromolecole della matrice extracellulare come l'elastina (Malech et al., 1987). I macrofagi possono anche essere coinvolti nel rimodellamento delle vie aeree tramite la secrezione di fattori di crescita come quello derivato dalle piastrine (PDGF), il fattore di crescita basico dei fibroblasti (b-FGF) e il fattore di crescita trasformante (TGF-β) (Vignola et al., 1996).
2 Mediatori chimici
Chemochine. Le chemochine sono importanti nel reclutamento delle cellule infiammatorie nelle vie aeree e sono espresse prevalentemente nelle cellule epiteliali (Wenzel, 2003). L'eotassina è relativamente selettiva per gli eosinofili, mentre le chemochine correlate all'attivazione timica (TARC) e le chemochine derivate da macrofagi (MDC) reclutano le cellule Th2.
Cistenil-leucotrieni. I cistenil-leucotrieni sono potenti broncocoscrittori principalmente derivati dai mastociti e la loro inibizione farmacologica comporta un notevole beneficio clinico nell'asma che consiste in un miglioramento della funzione polmonare e della sintomatologia asmatica (Miller et al., 2004). Citochine. Le citochine più importanti comprendono l'IL-1β e il TNF-α, che amplificano la risposta infiammatoria, ed il GM-CSF che prolunga la sopravvivenza degli eosinofili. Le citochine derivate dai linfociti Th2 includono IL-5, necessaria per la differenziazione e la sopravvivenza degli eosinofili; IL-4, importante per la differenziazione delle cellule Th2, IL-13, necessaria per formazione di IgE.
Istamina. E' un mediatore chimico liberato dai mastociti che contribuisce alla broncocostrizione ed alla risposta infiammatoria.
Ossido Nitrico (NO). E' una molecola prodotta principalmente dall'azione dell'enzima ossido nitrico sintetasi inducibile delle cellule epiteliali. L'NO nell'esalato viene valutato, in quanto marker associato con la presenza d'infiammazione nell'asma, per monitorare l'efficacia del trattamento antiasmatico (Smith et al., 2005).
Prostaglandina D2. E' una prostaglandina ad attività broncocostrittrice prodotta principalmente dai mastociti e coinvolta nel reclutamento delle cellule Th2 nelle vie aeree.
3 Le microparticelle
Le microparticelle (MP) costituiscono una popolazione eterogenea di vescicole rivestite di membrana, che possono essere rilasciate, da tutti i tipi di cellule durante l'attivazione o l'apoptosi, nel sangue e negli altri fluidi biologici. Furono identificate per la prima volta nel 1967 da Wolf che aveva dimostrato la presenza nel plasma umano di piccoli frammenti derivati da piastrine. Studi successivi hanno dimostrato che queste vescicole si possono formare sia da cellule normali (macrofagi, monociti, linfociti B e T, neutrofili, eritrociti, cellule endoteliali, cellule muscolari lisce vascolari e cellule epiteliali) che tumorali. A lungo considerate sottoprodotti cellulari inerti, le MP sono ora considerate elementi subcellulari funzionalmente attivi ben distinti da altre strutture subcellulari come gli esosomi e i corpi apoptotici, coinvolte in numerosi processi fisiopatologici, come la coagulazione del sangue e l'infiammazione.
In vitro, il rilascio delle microparticelle può essere indotto in diversi tipi di cellule e la composizione delle microparticelle cambia in funzione dello stimolo e delle linee cellulari di origine. Le MP possono svolgere un'azione autocrina o paracrina ed esercitare i loro effetti a distanza dal loro sito di origine. Sono presenti nel sangue periferico degli individui sani, ma i loro livelli aumentano in pazienti con malattie infiammatorie autoimmuni e aterosclerotiche (Ardoin, 2007).
Composizione delle microparticelle
Le MP (Figura 3) sono microvescicole di 0.05-1 µm, formate da una membrana che ha la struttura del plasmalemma della cellula da cui originano oltre ai contenuti citoplasmatici (Ardoin, 2007). Sono costituite prevalentemente da lipidi e proteine. La parte lipidica consiste in un doppio strato fosfolipidico che varia in base al tipo di cellula da cui derivano, o in base allo stimolo che le ha indotte (Morel, 2005); infatti, è stato visto che la percentuale di fosfolipidi che compone il doppio strato varia tra individui sani e pazienti (Boulanger, 2001).
Per quanto riguarda la parte proteica, le MP presentano le proteine di superficie della cellula da cui provengo. Le MP presentano una vasta gamma di molecole stimolatrici quali i recettori delle citochine, oltre ad acido arachidonico e DNA (Distler, 2006). Poiché hanno una composizione lipidica e proteica assai varia, esse costituiscono una fonte concentrata ed eterogenea di molecole che può svolgere un ruolo nella regolazione di molti processi biologici (Distler, 2006).

Fig. . Struttura della Microparticella
Generazione delle MP
I meccanismi intracellulari che portano alla liberazione delle MP non sono stati ancora completamente chiariti; è certo, comunque, che le MP vengono liberate dalle cellule nel corso di due processi biologici distinti: l'attivazione cellulare e l'apoptosi.
In vitro, l'attivazione cellulare si può osservare in seguito a stimolazione con diversi agonisti quali la trombina, il calcio ionoforo A23187 e lipopolisaccaridi (LPS), che determinano un aumento di Ca + citosolico, specialmente nel sito di vescicolazione. Infatti, durante l'attivazione cellulare si rileva un aumento della concentrazione intracellulare di Ca++, dovuto al rilascio di questo ione dai compartimenti intracellulari e al suo ingresso dall'ambiente extracellulare, con conseguente distacco della membrana plasmatica dal citoscheletro. Il distacco è causato dall'attivazione della calpaina, una proteasi che degrada le proteine di legame talina e actinina. La via della calpaina però non può essere l'unico meccanismo calcio dipendente per il rilascio delle microparticelle dal momento che la calpaina è inibita dalla calpeptina ma l'azione di quest'ultima non blocca anche il rilascio delle MP (Distler, 2006). Con l'attivazione cellulare, all'aumento di calcio segue un rimodellamento della membrana plasmatica, con modifiche che includono l'esposizione sul versante esterno della fosfatidilserina e la formazione di vescicole, ovvero MP, che portano con se proteine di superficie e componenti citoplasmatiche della cellula di origine.
La membrana plasmatica cellulare è formata da due strati fosfolipidici che hanno una particolare composizione. I due strati sono disposti in modo da esporre verso l'esterno della cellula e verso il citoplasma cellulare le teste idrofile dei fosfolipidi e verso l'interno della membrana le code idrofobe. Gli aminofosfolipidi (fosfatidilserina e fosfatidiletanolammina) sono posizionati specificatamente sul versante interno della membrana, mentre la fosfatidilcolina e la sfingomielina sono concentrati verso l'esterno della cellula. La distribuzione dei lipidi di membrana è controllata da tre diversi meccanismi:
. il primo meccanismo di controllo consiste in una pompa specifica per la fosfatidilserina e la fosfatidiletanolammina diretta verso l'interno (flippasi) conosciuta come aminofosfolipidetraslocasi. Questa pompa è fisiologicamente attiva; infatti la maggior concentrazione di fosfatidilserina è presente nel versante interno della membrana.
. il secondo meccanismo di controllo è una pompa che porta i fosfolipidi verso l'esterno della cellula (floppasi).
. il terzo meccanismo di controllo consiste in uno scambiatore di lipidi che promuove lo scambio bidirezionale non specifico dei fosfolipidi attraverso la membrana.
Un aumento significativo e sostenuto del Ca + intracellulare può condurre al collasso della membrana in maniera asimmetrica attivando lo scambiatore dei lipidi e la floppasi, e inibendo contemporaneamente l'attività delle flippasi. Il cambiamento più evidente nella distribuzione lipidica è l'esposizione delle fosfatidilserine dovuto all'inibizione delle flippasi, seguita poi dalla degradazione proteolitica del citoscheletro Ca -dipendente e dal rilascio delle microparticelle (Figura 4).
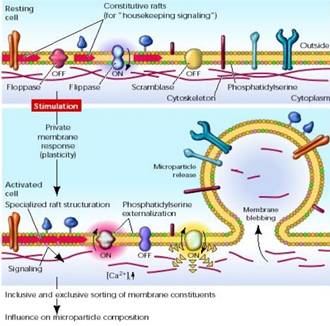
Fig. . Meccanismo di rilascio delle micro particelle
Per quanto riguarda il loro rilascio durante l'apoptosi, questo è associato al fenomeno del blebbing di membrana, ovvero al processo che vede la formazione di estroflessioni superficiali (blebs) seguita dalla frammentazione in corpi apoptotici. L'apoptosi è una forma di morte cellulare il cui fine è l'eliminazione di cellule non desiderate dell'organismo attraverso l'attivazione di una sequenza di eventi coordinati e programmati internamente alla cellula, messi in atto da una serie di prodotti genici specializzati. Si verifica per esempio durante lo sviluppo, come meccanismo omeostatico di mantenimento delle popolazioni cellulari all'interno di un tessuto, come meccanismo di difesa nelle reazioni immunitarie, o nell'invecchiamento. Le cellule che vanno incontro a questo fenomeno presentano le seguenti caratteristiche: diminuzione delle dimensioni cellulari, condensazione della cromatina e la formazione di protuberanze ("blebs") citoplasmatiche e di corpi apoptotici (Robbins, 1999). Il rilascio delle MP sembra avvenire tardivamente nell'apoptosi, contemporaneamente alla frammentazione cellulare e alla formazione dei corpi apoptotici. Il blebbing di membrana durante l'apoptosi dipende dall'attivazione di ROCK1, una chinasi-1 associata a Rho (Ardoin, 2007). Le proteine ROCK sono attivate dal legame di Rho al GTP e sono importanti mediatori per la riorganizzazione del citoscheletro. In esperimenti con fibroblasti di topo NIH3T3 e cellule epiteliali umane di tumore della mammella MCF10A la vescicolazione delle MP è diminuita per il blocco della ROCK 1 da parte di una piccola molecola inibitrice Y27632 (Distler, 2006). Probabilmente a seconda che le MP vengano prodotte in seguito ad attivazione o apoptosi variano sia i meccanismi con cui si formano che le dimensioni, la composizione interna e di superficie; infatti l'espressione di molecole di membrana sulla superficie delle MP di derivazione endoteliale varia a seconda che la loro formazione risulti dall'attivazione cellulare o dall'apoptosi. Non è ancora noto se queste differenze strutturali influenzino anche la loro funzione (Ardoin, 2007).
Le microparticelle possono inoltre essere prodotte da cellule necrotiche dopo perdita dell'integrità della membrana o dopo distruzione meccanica delle cellule stesse in quanto viene persa l'asimmetria della distribuzione dei fosfolipidi. La fosfatidilserina, una volta accessibile alla cellula o alle microparticelle, è importante perché regola due funzioni fisiologiche importanti:
stimola la coagulazione del sangue.
costituisce il sistema di riconoscimento delle cellule senescenti da parte del sistema reticolo endoteliale.
Non tutti i lipidi e le proteine però hanno la stessa mobilità all'interno della membrana cellulare come evidenziato durante il clustering dei recettori, la dimerizzazione e il capping dei leucociti. Studi recenti hanno dimostrato che i lipidi di membrana e le proteine sono organizzati in domini raft, cioè sono come isole mobili di determinata composizione lipidica e proteica disposte sulla membrana (Horstman et al., 2004). L'organizzazione in domini raft fornisce un razionale che spiega il rilascio di particolari lipidi e proteine con la formazione delle microparticelle (Hugel et al., 2004).
Funzioni delle MP
In contrasto con la loro iniziale definizione di detriti cellulari inerti, le MP attualmente sono considerate molecole fisiologicamente attive coinvolte in almeno tre importanti processi biologici: l'infiammazione, la coagulazione e la funzione vascolare (Ardoin, 2007).
Sono state fatte molte ipotesi sulla funzione delle MP:
Comunicazione intracellulare e intercellulare. I primi organismi unicellulari (ad esempio, le amebe) erano in grado di produrre MP per comunicare con gli organismi vicini.
Vettori virali o vettori metastatizzanti.
Meccanismo di resistenza ai farmaci in cellule tumorali
Mediatori dell'infiammazione.
Inoltre le MP possono essere utilizzate come potenziali marker patologici poiché studi recenti hanno dimostrato che esiste una correlazione tra l'aumento del numero di MP derivanti da determinate cellule e di conseguenza aventi particolari caratteristiche fenotipiche ed alcune patologie, come ad esempio l'artrite reumatoide ed il lupus (Sellam et al., 2009)
Le microparticelle sono quindi coinvolte in molti processi biologici quali la coagulazione (Celi et al., 2004) e l'infiammazione (Gasser et al., 2003). Il ruolo delle microparticelle nelle malattie infiammatorie quali l'artrite reumatoide, è stato oggetto di studi recenti ed è stato indagato anche il loro ruolo nel rilascio di citochine dai leucociti (Distler et al., 2005) e da cellule epiteliali polmonari (Cerri et al., 2006).
L'epitelio delle vie aeree partecipa attivamente nella difesa contro i patogeni a contatto con le vie aeree superiori e regola le reazioni infiammatorie che caratterizzano le malattie respiratorie. Il reclutamento di leucociti circolanti nel sangue periferico rappresenta una fase cruciale in questi processi e richiede l'azione combinata di chemochine e molecole di adesione; le MP hanno un ruolo anche in questo ambito.
L'implicazione delle microparticelle nel processo infiammatorio e l'elevata concentrazione riscontrata nelle vie aeree dei pazienti asmatici suggeriscono che le microparticelle svolgano un ruolo centrale nella patogenesi delle malattie respiratorie. Il meccanismo d'azione delle microparticelle nel sistema respiratorio e nel sistema immunitario rappresenta quindi un argomento di particolare interesse scientifico per comprendere meglio i processi molecolari legati all'infiammazione e promuovere lo sviluppo di nuove strategie terapeutiche nelle malattie croniche respiratorie.
|
| Appunti su: secrezione di citochine infiammatorie nelle vie respiratorie, |
|
| Appunti Bellezza |  |
| Tesine Bambini |  |
| Lezioni Nutrizione |  |