|
| Appunti superiori |
|
|
| Appunti superiori |
|
| Visite: 2995 | Gradito: |
Leggi anche appunti:Biologia molecolare del muscoloBiologia molecolare del muscolo Esistono tre tipi di muscolo: Apparato digerenteApparato digerente 1 INTRODUZIONE Il fascino della gola del furloIL FASCINO DELLA GOLA DEL FURLO Nella penombra e nel silenzio che regna |
 |
 |
Comparse forse già nel Triassico (circa 200 milioni di anni fa) o più probabilmente nel successivo Giurassico, le angiosperme si diffusero rapidamente in un grande numero di forme diverse a partire dalla fine del Cretaceo (90 milioni di anni fa circa). Molti aspetti dell'origine delle piante a fiore sono ancora misteriosi. Non si sa con precisione né quando e dove si siano originate, né quale gruppo di gimnosperme ne sia il progenitore e neppure se si tratti di un gruppo monofiletico oppure no. Anche se praticamente tutti i gruppi di gimnosperme conosciuti allo stato fossile sono stati di volta in volta indicati dai diversi autori come possibili antenati diretti delle piante a fiore, appare abbastanza convincente l'ipotesi di un'affinità filogenetica delle angiosperme con la linea che ha portato anche alle Cycadopsida, soprattutto se si considera la somiglianza tra i macrosporofilli fogliacei con ovuli marginali presenti ancora oggi in alcuni rappresentanti di questo gruppo di gimnosperme e il macrosporofillo ancestrale che ripiegandosi lungo la nervatura centrale avrebbe dato origine al carpello delle angiosperme. Non è chiaro nemmeno come abbiano fatto le piante a fiore a diversificarsi così rapidamente. C'è chi ritiene che questo possa essere spiegato con un'origine in zone montuose tropicali con climi caratterizzati da alternanza di periodi umidi e aridi, zone cioè dove sono presenti forti pressioni selettive che avrebbero provocato una rapida evoluzione e insieme però anche condizioni sfavorevoli alla formazione e conservazione dei fossili. Le prime tappe della comparsa e della diversificazione delle piante a fiore si sarebbero quindi svolte senza lasciare tracce. Sta di fatto che in un tempo relativamente breve le angiosperme sono diventate il gruppo di vegetali che domina il nostro pianeta, grazie alla loro estrema varietà morfologica e fisiologica e in particolare alla diversità di tipi di foglie e di tessuti conduttori, che consentono loro la vita negli ambienti più diversi. Il risultato è che le Magnoliophyta sono i vegetali dominanti in tutti i grandi biomi terrestri, escluse la tundra e la foresta di conifere e costituiscono un elemento fondamentale del paesaggio, naturale o modificato dall'uomo. Con circa 250.000 specie viventi, riunite in circa 400 famiglie (ma il numero varia di molto a seconda degli autori), sono il gruppo di piante più numeroso e vario: basti pensare che tutti gli altri gruppi di tracheofite (pteridofite e gimnosperme) comprendono insieme solo 15.000 specie circa. Le angiosperme hanno anche un'enorme importanza applicativa ed economica, comprendendo tutte le principali piante coltivate (a scopo alimentare, medicinale, foraggero, ornamentale, tessile, ecc.), nonché la maggior parte delle infestanti.
Le angiosperme sono caratterizzate da un'impressionante varietà di forme, a cominciare dalla fisionomia generale della pianta: si va da alberi giganteschi a erbe annue, a arbusti, liane, piante epifite, succulente, insettivore, parassite prive di clorofilla, ecc. Forme diversissime sono presenti in tutti gli organi della pianta, in particolare nelle foglie e nei fiori, così da consentire lo svolgimento delle fondamentali funzioni vegetative e riproduttive negli ambienti più vari. La struttura secondaria del fusto è caratterizzata dalla presenza di legno di tipo eteroxilo, dove elementi specializzati per le diverse funzioni di conduzione e sostegno (trachee, tracheidi, fibre) sono fra loro combinati nelle proporzioni più adatte a garantire la miglior efficienza del tessuto nelle diverse condizioni ambientali.[2]
Una tale diversità e capacità di adattamento è stata raggiunta dalle piante a fiore grazie soprattutto alla grande possibilità di insorgenza di nuovi caratteri legata all'affinamento dei processi riproduttivi, che raggiungono qui la massima efficienza e perfezione nell'ambito dei vegetali.
Il fiore e gli elementi fiorali.
Caratteristica del gruppo è la presenza del carpello, che non è altro che un macrosporofillo entro cui sono contenuti e racchiusi gli ovuli (macrosporangi rivestiti da tegumenti). Il carpello dà luogo a una struttura detta pistillo, formata in genere da tre porzioni con funzioni diverse:
l'ovario, parte allargata basale che contiene gli ovuli;
lo stimma, parte apicale recettiva per il polline;
lo stilo, parte allungata che collega l'ovario allo stimma.
Il pistillo (ovario-stilo-stimma) è un'unità morfologico-funzionale che può essere formata da un solo carpello nel caso di ginecei monocarpellari o pluricarpellari apocarpici (e in questo caso i termini carpello e pistillo coincidono) o da più carpelli fusi insieme nel caso di ginecei pluricarpellari sincarpici.
I carpelli, insieme alle altre strutture riproduttive, sono organizzati nel fiore, il complicato e perfezionatissimo organo esclusivo delle angiosperme[3] da qualcuno definito come un insieme di strutture che proteggono e nutrono i gametofiti e controllano la gamia (Gerola).
Il fiore è un complesso specializzato in cui sono riuniti ed organizzati tutti gli organi e le strutture legati alla riproduzione. È qui che:
- si formano i macro- e i microsporangi;
- avviene la meiosi, con produzione delle macro- e delle microspore;
- vengono prodotti, nutriti e protetti i gametofiti maschile e femminile;
- vengono messi in atto i processi di incompatibilità controllati geneticamente che favoriscono il raggiungimento dell'oosfera da parte del gamete maschile "più adatto";
- avviene la gamia e il successivo sviluppo dell'embrione, primo stadio vitale del nuovo sporofito;
- attraverso la formazione del seme e del frutto, vengono forniti al nuovo sporofito i mezzi per affrontare con le maggiori probabilità di successo il distacco dalla pianta madre e la diffusione in un nuovo ambiente.
Il fiore è un germoglio a crescita determinata con internodi raccorciati e nodi che portano foglie specializzate: antofilli sterili (sepali e petali, che costituiscono il perianzio) e sporofilli fertili (microsporofilli o stami e macrosporofilli o carpelli).
L'enorme varietà morfologica e biologica che si trova nei fiori è il risultato di un lungo processo evolutivo nel corso del quale la selezione ha premiato quelle caratteristiche che nei diversi ambienti si sono rivelate più adatte a favorire l'efficienza della riproduzione. Si ritiene che nei fiori delle prime angiosperme gli elementi fiorali fossero in numero elevato e indefinito, disposti a spirale su un asse (ricettacolo o talamo) ben sviluppato. Fiori di questo tipo si ritrovano ancora in angiosperme che hanno conservato caratteri di primitività, come le magnolie o le ninfee.
È possibile individuare alcune tendenze evolutive che si sono ripetute molte volte indipendentemente nell'evoluzione dei fiori dei diversi gruppi di angiosperme, come ad esempio:
- da fiori con elementi numerosi a fiori con elementi in numero ridotto;
- da fiori con numero variabile di pezzi a fiori con numero fisso di elementi per ogni specie;
- da disposizione spiralata dei pezzi fiorali (fiori aciclici) a disposizione in verticilli, dapprima solo di una parte degli elementi fiorali (fiori emiciclici, come nella fragola) e poi di tutti (fiori ciclici, come nella maggioranza delle angiosperme attuali);ù
- da fiori con elementi tutti liberi (calice dialisepalo, corolla dialipetala, gineceo apocarpico, ecc.) a fiori con elementi saldati nello stesso verticillo (calice gamosepalo, corolla gamopetala, gineceo sincarpico, ecc.) o tra verticilli contigui (ad esempio stami epicorollini, ovario infero);
- da fiori a simmetria radiata a fiori zigomorf;
- da fiori in cui ssono presenti tutti i verticilli a fiori in cui uno o più mancano..
I verticilli di un fiore completo sono (dal basso verso l'alto se il ricettacolo è allungato ed evidente, o dall'esterno verso l'interno nel caso più comune di ricettacoli brevi):
- calice, formato da sepali, con funzione di protezione degli altri verticilli nelle prime fasi del loro sviluppo
- corolla, formata da petali, prevalentemente con funzione di richiamo sul fiore degli insetti impollinatori. L'origine dei petali non è la stessa in tutte le angiosperme: in alcuni casi si tratta chiaramente di sepali modificati, in altri appare evidente la loro origine da stami sterili sovrannumerari. L'insieme di calice e corolla forma il perianzio. Se i due verticilli perianziali non sono molto diversi per aspetto e funzione, prendono il nome di tepali e il loro insieme di perigonio. Il perigonio può avere aspetto corollino (tepali petaloidi) o calicino (tepali sepaloidi). La presenza di un perigonio anziché di un perianzio è la condizione più frequente nelle monocotiledoni.
- androceo (di solito in due verticilli), formato da microsporofilli o stami. Accanto a normali stami fertili (composti da filamento e antera) possonoe ssere presenti stami sterili (staminodi) più o menio modificati, con funzioni diverse, spesso di richiamo per gli insetti pronubi in aggiunta o in sostituzione dei petali;
- gineceo, formato da macrosporofilli o carpelli. Fiori dove sono presenti sia stami che carpelli si chiamano perfetti (o monoclini o ermafroditi). I fiori diclini (o unisessuali) portano invece solo stami (fiori maschili) o solo carpelli (fiori femminili) e si trovano sullo stesso individuo nelle piante monoiche (come il mais) e su individui diversi nelle piante dioiche (come i salici).
Non sempre tutti i verticilli sono presenti. Uno o più di essi possono mancare e si possono così avere fiori monoclamidati, fiori nudi, fiori unisessuali, fiori sterili, ecc. La successione dei verticilli è però la stessa in tutte le angiosperme e questo costituisce un indizio a favore di una loro origine monofiletica. Nei fiori impollinati da insetti è frequente la presenza di nettàrii, strutture più o meno evidenti dove viene raccolto il nèttare. I nettàrii possono trovarsi alla base dei petali, sul ricettacolo, sulle pareti o alla base dell'ovario o altrove; anche petali o sepali possono essere trasformati parzialmente o totalmente in nettàrii.
La disposizione verticillata ha consentito la fusione tra elementi di ciascun verticillo o di verticilli adiacenti, con la comparsa - ad esempio - di calici gamosepali e corolle gamopetale, di stami epicorollini, del passaggio alla posizione infera dell'ovario con la saldatura sulla sua parete delle basi degli altri pezzi fiorali.
Nella maggior parte dei casi, la selezione naturale ha premiato i tipi di fiori in cui viene favorita l'allogamia (cioè l'unione tra gameti geneticamente diversi perché provenienti da fiori o addirittura da individui diversi), che offre maggiori possibilità di comparsa di nuovi caratteri. Ciò non toglie che in alcuni casi ci sia stata invece un'evoluzione verso l'autogamia (o addirittura verso forme diverse e anche molto raffinate di propagazione vegetativa, come l'apomissia), laddove le condizioni ambientali la rendevano vantaggiosa.
Molte caratteristiche del fiore sono legate al tipo di impollinazione, cioè di trasporto del polline dalle antere degli stami fino allo stimma del pistillo. Nei fiori entomogami (o entomofili), in cui il trasporto del granulo è affidato a insetti, si ha in genere un particolare sviluppo della corolla, con forme, colori, odori, strutture meccaniche atte a favorire la visita degli animali, massimizzando l'efficienza nel trasporto del polline, minimizzando i danni e nelle forme più evolute arrivando a selezionare i pronubi. La varietà dei fiori entomogami è il risultato di processi di coevoluzione che hanno interessato angiosperme e insetti dal Cretaceo in avanti. Al contrario, nei tipici fiori anemogami, dove è il vento il vettore del polline, si assiste ad un insieme di adattamenti morfo-funzionali che facilitano il trasporto, da alcuni indicati come "sindrome anemogama": polline abbondante e leggero, stami con antere mobili e filamenti lunghi, perianzio ridotto o assente e privo di profumi, stimmi voluminosi e piumosi, elevato rapporto numero di stimmi/numero di ovuli, da cui frutti spesso monospermi. L'anemogamia è diffusa tra le specie erbacee di ambienti aperti (praterie), come molte graminacee e tra le specie arboree degli strati superiori delle foreste. Un tempo le angiosperme anemofile, caratterizzate da fiori con struttura semplice e ridotta, erano considerate meno evolute rispetto a quelle con i più complessi fiori entomogami, secondo un'interpretazione che vedeva l'evoluzione come un processo diretto verso una sempre maggiore complessità delle strutture. In seguito si sono trovate le prove che angiosperme anemofile a fiori estremamente ridotti erano derivate da forme con fiori entomofili complessi. È bene tenere sempre presente che non esistono caratteri in assoluto più e meno evoluti, ma solo più e meno adatti a particolare condizioni ambientali che li rendono vincenti nella selezione. Un carattere punito dalla selezione in un ambiente e in un'epoca può essere premiato in un ambiente diverso o ricomparire in un'epoca successiva.
La riproduzione sessuale.
Come detto, nel fiore sono riunite le strutture dove si svolgono tutte le fasi della riproduzione, fasi che nelle altre piante terrestri avvengono in luoghi diversi e al di fuori del diretto controllo dell'individuo genitore.
I microsporangi sono riuniti a gruppi di quattro a costituire le quattro logge o sacche polliniche delle antere degli stami (microsporofilli). In ogni sporangio si riconoscono, a partire dall'esterno, un tessuto protettivo bistratificato (esotecio e endotecio), un tappeto nutritivo e un tessuto sporigeno o archesporio, formato di cellule madri delle microspore, ognuna delle quali per meiosi originerà quattro microspore.
Le microspore, avvolte come nelle gimnosperme da esina e endina, germinano ancora nell'antera, producendo al loro interno un minuscolo gametofito costituito da una cellula vegetativa (o cellula del tubetto) che include una piccola cellula generativa. La maggior parte dei granuli pollinici (microspore germinate) lascia l'antera a questo stadio di sviluppo (pollini binucleati); in poche angiosperme più evolute, prima che il polline venga rilasciato la cellula generativa si divide in due gameti maschili o cellule spermatiche (pollini trinucleati). Le caratteristiche morfologiche dell'esina e in particolare la presenza e il tipo di solchi o pori di germinazione sono specifiche nei diversi gruppi sistematici e possono consentire l'identificazione comunemente fino al livello della famiglia e in qualche caso fino al livello di specie.
Trasportati dagli agenti dell'impollinazione (vento, insetti o altro), i granuli raggiungono lo stimma. Se il polline è di tipo compatibile, lo stimma secerne zuccheri, sali minerali, vitamine e altre sostanze che stimolano l'idratazione del granulo, provocando l'aumento di volume del citoplasma e la fuoriuscita del tubetto pollinico. Questo si fa strada nei tessuti lassi dello stilo, secernendo esoenzimi (pectinasi) che solubilizzano le lamelle mediane delle cellule. Nei pollini binucleati è durante la crescita del tubetto che avviene la formazione dei due gameti maschili.
Nel frattempo, all'interno degli ovuli contenuti negli ovari, si è prodotto il ridottissimo gametofito femminile o sacco embrionale. Nella nucella dell'ovulo una sola cellula madre delle macrospore subisce meiosi e origina quattro spore, di cui di regola una sola resta vitale, mentre le tre più prossime al micropilo degenerano. Il nucleo della macrospora si divide in due nuclei che migrano ai poli opposti della cellula, sospinti da un grosso vacuolo centrale. Con due successive divisioni di ciascun nucleo, si ottengono quattro nuclei al polo micropilare e quattro al polo opposto (calazale). Due nuclei, uno per ciascun polo, si trasferiscono al centro del sacco embrionale e si rivestono di una parete comune (nuclei polari). Anche gli altri sei nuclei si cellularizzano, completando così lo sviluppo del gametofito, che nel tipo più comune è costituito da sette sole cellule: l'oosfera accompagnata da due sinergidi al polo micropilare, tre antipodi al polo calazale e una cellula con due nuclei migrati dai due poli (nuclei polari) al centro. Il sacco embrionale è ora pronto per la fecondazione.
I gameti maschili, portati dal tubetto, sono frattanto giunti fino al gametofito femminile, dove il tubetto si apre riversandoli di solito in una delle due sinergidi. Da qui una delle due cellule spermatiche raggiunge l'oosfera, a cui si unisce per formare lo zigote, mentre l'altra si fonde con i nuclei polari, originando l'endosperma secondario, tessuto di riserva normalmente triploide tipico delle angiosperme. In alcune specie il tessuto è subito cellulare, in altre passa prima per uno stadio a nuclei liberi. Il processo di doppia fecondazione, che avviene in tutte le angiosperme con modalità simili, è uno degli argomenti a sostegno dell'ipotesi di un'origine monofiletica delle Magnoliophyta. A differenza dell'endosperma primario delle gimnosperme, l'endosperma secondario si forma solo in seguito all'avvenuta fecondazione, realizzando il massimo di efficienza e di risparmio energetico. Nelle gimnosperme invece l'endosperma primario si forma indipendentemente dalla fecondazione: in quelle più arcaiche prima ancora dell'arrivo del polline, in quelle più evolute (Pinopsida) in genere in seguito all'impollinazione.
Dopo la fecondazione, l'ovulo si trasforma in seme. Dallo zigote si sviluppa un embrione, spinto dalla sua parte basale (sospensore) dentro l'endosperma. A differenza di quanto avviene nelle gimnopsperme, l'embrione delle angiosperme è cellularizzato fin dall'inizio dello sviluppo, come nelle pteridofite. A un certo punto del suo sviluppo, che si svolge con modalità diverse nelle dicotiledoni e nelle monocotiledoni, l'embrione arresta la crescita. In questo stadio sono in genere riconoscibili uno o due foglie embrionali (cotiledoni) che circondano un apice caulino (piumetta), e una radichetta embrionale con il suo apice meristematico. A raccordare cotiledoni e radichetta può essere visibile un asse ipocotile, più sviluppato nelle specie a germinazione epigea. Attorno all'embrione si organizza l'endosperma, dapprima in forma nucleare e poi cellularizzato. A spese dello sporofito genitore, l'endosperma si arricchisce di sostanze di riserva, diverse a seconda delle specie: amidi (come nei cereali), proteine (legumi), lipidi (ricino, olivo), emicellulose, e altro. In molte dicotiledoni, nel corso dello sviluppo del seme le riserve vengono trasferite interamente ai cotiledoni e l'endosperma viene completamente riassorbito e non è presente nel seme maturo. Mentre si formano l'embrione e le riserve, i tegumenti dell'ovulo si modificano, a costituire i tegumenti del seme. Quando questo ha terminato il suo sviluppo, va incontro ad un processo di disidratazione, mentre l'embrione entra in quiescenza. Tornerà a idratarsi e riprenderà la crescita solo quando, lontano nel tempo e nello spazio dalla pianta madre, troverà l'ambiente adatto alla germinazione e alla successiva crescita della nuova pianta. Ad impedire che questo avvenga prima che tutte le condizioni ambientali siano favorevoli, in molti semi di angiosperme sono presenti fenomeni di dormienza che non consentono la germinazione prima che siano rimossi blocchi di tipo meccanico o biochimico.
Anche i semi delle angiosperme, come avviene in alcune gimnosperme, possono presentare strutture che ne facilitino la dispersione, come ali (betulla) o filamenti (pioppi) nelle specie disseminate dal vento, o porzioni appetibili (arilli, caruncole, strofioli) in quelle che sfruttano gli animali. Ma nelle piante a fiore questo compito viene svolto anche e soprattutto dal frutto, organo specializzato esclusivo delle angiosperme che deriva dalla trasformazione del pistillo - e in particolare delle pareti dell'ovario - dopo la fecondazione. Essendo null'altro che una foglia modificata, l'ovario avrà una struttura anatomica a tre strati, che corrispondono all'epidermide superiore e inferiore e al mesofillo. Questi nel frutto vanno a costituire il pericarpo, in cui spesso è possibile individuare tre diversi tessuti: epicarpo, mesocarpo e endocarpo. Il pericarpo si arricchisce di acqua e zuccheri nei frutti carnosi che per la disseminazione devono essere ingeriti dagli animali e assume consistenza cartacea o legnosa nei frutti secchi. I tanti tipi di frutti che si possono osservare non sono altro che il risultato delle possibili modificazioni dei diversi tipi di gineceo (mono- o pluricarpellare, apocarpico o sincarpico, con ovario supero o infero, ecc.) in funzione dei diversi agenti di dispersione.Tra i frutti carnosi, la drupa (pesca, oliva) deriva da ovari che contengono un solo seme ed è caratterizzata da avere un endocarpo legnoso; la bacca (pomodoro, uva) deriva in genere da ovari pluricarpellari sincarpici contenenti più semi. Tipi particolari di bacca sono stati descritti con nomi particolari: ad esempio il peponide è un tipo di bacca con epicarpo duro a maturità, caratteristico della famiglia delle cucurbitacee (zucca, melone). I frutti secchi vengono in genere distinti in frutti deiscenti (che si aprono a maturità per far fuoriuscire i semi) e frutti indeiscenti. I frutti secchi indeiscenti sono di norma monospermi, cioè contengono un solo seme e hanno l'aspetto di semi piuttosto che di frutti, tanto che molti vengono considerati comunemente semi (ad esempio un "seme" di girasole, in realtà un achenio). Il tipo più semplice di frutto secco indeiscente è l'achenio. Gli altri possono essere considerati modificazioni di acheni: la sàmara (frassino, olmo) è un achenio con pericarpo espanso in un'ala che facilita la dispersione ad opera del vento; la cariosside (graminacee) è un achenio con pericarpo che si salda sui tegumenti del seme. Alcuni ovari pluricarpellari si suddividono a maturità in porzioni contenenti un solo seme, ciascuna con l'aspetto di un singolo frutto secco indeiscente (schizocarpi): così la disàmara degli aceri che si divide in due sàmare, il tetrachenio delle labiate che si separa in quattro acheni. I frutti secchi deiscenti contengono in genere più semi e a maturità si aprono per consentire a ciascun seme di diffondersi autonomamente. Comprendono il follicolo (elleboro, oleandro), che si apre lungo una sola linea di deiscenza; il legume (pisello, robinia), che deriva da un gineceo monocarpellare e a maturità si apre lungo due linee, corrispondenti alla nervatura centrale e alla linea di sutura dei margini del macrosporofillo; la siliqua (senape, rapa), che deriva da un ovario bicarpellare sincarpico e si apre in due valve che lasciano sul peduncolo fiorale un setto su cui sono portati i semi; la capsula (iris, paulownia), che deriva da ovari pluricarpellari sincarpici e contiene in genere numerosi semi. Le capsule possono aprirsi con modalità diverse (per setti, pori, coperchi) e tipi particolari di capsule sono indicati con nomi particolari: ad esempio il treto del papavero, tipo di capsula che libera i semi attraverso pori apicali. Da ginecei pluricarpellari apocarpici si possono originare frutti composti (o frutti aggregati), come nel caso della mora di rovo o del lampone; da infiorescenze derivano invece le infruttescenze dell'ananas o del gelso, dove gli ovari dei diversi fiori che compongono l'infiorescenza si saldano fra loro a formare un'unica struttura. Vengono considerati falsi frutti quei frutti in cui la parte carnosa non è data dalla trasformazione delle pareti dell'ovario, ma da altri organi: nella fragola la porzione rossa e zuccherina deriva dall'ingrossamento del ricettacolo su cui erano inseriti i numerosi carpelli monospermi che a maturità si trasformano negli acheni disseminati sulla superficie della fragola; nel pomo (mela, pera) che deriva da ovario infero, le pareti dell'ovario corrispondono grosso modo al torsolo, mentre la parte carnosa deriva dall'ingrossamento dei tessuti dell'ipanzio[4].
La dispersione può avvenire ad opera di vari agenti e può interessare i singoli semi, i frutti, o entrambi: gli animali trasportano a distanza frutti o semi delle specie zoocore, che vengono ingeriti e poi espulsi successivamente o aderiscono al vello tramite aculei o altre strutture; il vento è l'agente di dispersione delle angiosperme anemocore, che hanno frutti o semi leggeri, muniti di ali o altre appendici che facilitino il volo, come il pappo degli acheni delle composite o le brattee dei tigli; molte angiosperme di ambiente acquatico sono idrocore, cioè si servono dell'acqua per la dispersione e hanno semi o frutti adattati al galleggiamento e alla sopravvivenza in ambiente acquatico, come la palma da cocco. Poche specie autocore provvedono autonomamente alla dispersione, come il cocomero asinino (Ecballium elaterium), una cucurbitacea i cui frutti a maturità si aprono con violenza al minimo urto espellendo i semi, o numerose crucifere del genere Cardamine, le cui silique si aprono a scatto lanciando i semi a distanza. Semi o frutti privi di particolari adattamenti per la dispersione si allontanano dalla pianta madre grazie all'azione della forza di gravità (specie barocore).
La propagazione vegetativa spontanea è molto diffusa nei diversi gruppi di angiosperme. Il fondamento biologico della propagazione vegetativa sta nella proprietà delle cellule vegetali di mantenersi totipotenti non solo nella linea germinativa, ma anche in quella somatica, tanto che in opportune condizioni cellule di tessuti già differenziati possono essere indotte a sdifferenziarsi e tornare allo stato di tessuti meristematici. In questo modo, in condizioni favorevoli da tessuti di fusti possono svilupparsi radici avventizie. In natura si possono formare talee spontanee come adattamento a particolari condizioni ambientali, ad esempio in piante che crescono lungo corsi d'acqua (salici, oleandro). Molto diffusa tra le piante erbacee perenni è la propagazione spontanea per frammentazione di fusti modificati come stoloni, fusti rizomatosi, rizomi, tuberi, bulbi, ecc. È questo il mezzo di diffusione di molte infestanti (gramigna), che si avvantaggiano delle normali lavorazioni meccaniche che frammentano i fusti sotterranei. Molte angiosperme arboree sono in grado di emettere polloni dalla ceppaia in seguito al taglio, all'incendio o al morso di animali. Questa capacità viene sfruttata dall'uomo nel governo a ceduo dei boschi. Alcune angiosperme arboree particolarmente invadenti come la robinia e l'ailanto emettono polloni anche dalle radici.
In genere le piante capaci di propagazione vegetativa mantengono anche la capacità di riprodursi per seme. La propagazione vegetativa consente alla pianta una rapida diffusione in un ambiente uniforme, ma può risultare svantaggiosa in caso di cambiamento delle condizioni ambientali; la riproduzione sessuale interviene in quel caso a fornire nuovi genomi che possono rivelarsi più adatti alle condizioni mutate.
La propagazione vegetativa è da sempre utilizzata dall'uomo per mantenere inalterate nella discendenza le caratteristiche (organolettiche, di produttività, di resistenza) delle cultivar coltivate (cloni) e per ottenere in breve tempo molti individui già con caratteri adulti. Le tecniche di moltiplicazione più usate dall'uomo sono la talea, l'innesto e la propaggine e possono prevedere l'utilizzo di sostanze stimolanti (fitormoni del tipo delle auxine) e/o di particolari condizioni ambientali (serre di nebulizzazione). Attualmente sono molto utilizzate anche le tecniche di propagazione in vitro, che consentono di ottenere moltissime piantine in breve tempo a partire da pochissimo materiale. La tecnica prevede la coltura in ambiente sterile su mezzo nutritivo di frammenti di tessuti che vengono trattati con fitormoni: cambiando il rapporto citochinine/auxine viene stimolata dapprima la formazione di un tessuto indifferenziato (callo), poi l'insorgenza di germogli avventizi e infine di radici.
La sistematica delle Angiosperme
Secondo una visione tradizionale della sistematica basata essenzialmente sui caratteri morfologici, le angiosperme venivano divise nelle due classi delle monocotiledoni (Liliopsida) e dicotiledoni (Magnoliopsida). All'interno di queste ultime, venivano individuate in passato un piccolo numero di sottoclassi sulla base delle caratteristiche del perianzio: piante con fiori privi sia di calice che di corolla oppure provvisti di un solo verticillo; piante a fiori con corolla dialipetala; piante a fiori con petali fusi in corolla gamopetala. Tra queste sottoclassi veniva individuata una linea evolutiva che andava dal perianzio ridotto o assente, alla corolla a elementi liberi, a quella gamopetala. Sulla base di un'impostazione sistematica di questo tipo, tutte le angiosperme della flora mondiale vennero inquadrate dal sistematico tedesco Engler in uno schema tassonomico molto articolato che arrivava fino al rango di genere. Partendo dall'ordinamento di Engler, famiglie e generi sono stati ordinati in una sequenza sistematica su presunte basi filogenetiche, dal genere meno evoluto a quello più evoluto. Schematizzando molto, nelle dicotiledoni si andava dalle cosiddette amentifere - piante arboree con fiori anemogami a perianzio nullo o ridotto riuniti in amenti di famiglie come Salicaceae, Betulaceae, Fagaceae - fino alle più complesse composite (Asteraceae), con fiori a corolla gamopetala riuniti in infiorescenze specializzate a capolino. Nelle monocotiledoni, da famiglie con fiori regolari come Alismataceae, Juncaceae, Liliaceae, alle Orchidaceae con fiori irregolari complessi.
Questa impostazione sistematica ha goduto a lungo di un vasto seguito in gran parte d'Europa. Sequenze di famiglie e generi basate su un simile schema di riferimento sono tuttora alla base dell'ordinamento seguito in flore autorevoli anche recenti (Flora Europaea, Flora d'Italia di Pignatti) e nelle collezioni di molti erbari.
Tuttavia, indagini sistematiche condotte nell'ultimo mezzo secolo utilizzando altri dati oltre a quelli morfologici (dati anatomici, cromosomici e biosistematici in genere, biochimici) hanno messo in luce legami filogenetici e linee evolutive molto diversi da quelli ipotizzati da Engler e in particolare si è riconosciuto il carattere in genere derivato dei fiori anemofili rispetto a quelli entomofili. Sulla base di queste nuove conoscenze sono stati proposti vari schemi di classificazione delle piante a fiore di impostazione rigorosamente filogenetica, in molte delle quali scompare la stessa distinzione tra monocotiledoni e dicotiledoni.
Tra questi, uno dei più accettati è stato a lungo quello di Arthur Cronquist (in seguito integrato e modificato da altri autori), in cui le angiosperme più primitive non sono più ritenute le amentifere, ma quelle a fiori con elementi numerosi disposti a spirale, del tipo delle Magnoliaceae, Nymphaeaceae, Ranunculaceae, ecc. In questo schema, le piante a fiore sono divise in una decina di sottoclassi, il cui numero e la cui delimitazione hanno subìto via via numerose modifiche, col progredire delle conoscenze.
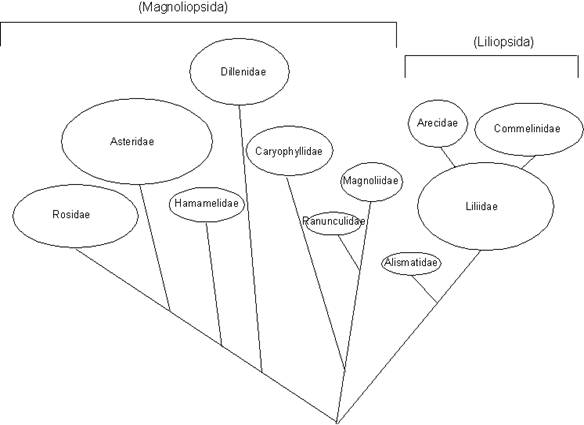
Fig. 5 - Una classificazione delle Magnoliophyta in 11 sottoclassi, basata sul sistema di Cronquist (modificato da Gerola Biologia e diversità dei vegetali Ed. UTET1995).
La classificazione di Cronquist è stata accettata da molti autori nei decenni passati, ma attualmente è ritenuta anch'essa superata sulla base delle nuove conoscenze derivate dall'analisi e confronto del DNA dei diversi taxa, attraverso la quale dovrebbe essere possibile ricostruire direttamente i legami filogenetici tra i diversi taxa. Questo tipo di indagini è sfociato in un nuovo approccio strumentale alla sistematica (sistematica molecolare), che ha portato a dare negli ultimi decenni nuovo impulso agli studi sulla filogenesi e conseguentemente sulla tassonomia delle angiosperme. Le indagini sistematiche sulle Magnoliophyta conoscono attualmente un momento di grande floridità, col risultato che nuovi schemi tassonomici vengono proposti si può dire ogni anno.
Al momento, nessuno di questi ha trovato l'accordo della maggioranza degli studiosi ed è accettato come risolutivo, tanto che in molti testi recenti le angiosperme vengono inquadrate in generi, famiglie, ordini, ma al di sopra di questo rango sono riunite in gruppi provvisori senza valore tassonomico.
In particolare, è stata messa fortemente in discussione la tradizionale distinzione delle angiosperme nelle due classi delle monocotiledoni e delle dicotiledoni (secondo le regole della nomenclatura botanica, meglio denominate come classi Liliopsida e Magnoliopsida). Inizialmente si è ritenuto che la separazione tra le due classi fosse avvenuta molto precocemente, dal momento che tra i resti fossili del Cretaceo si possono riconoscere già forme attribuibili all'una o all'altra e che alcune famiglie di angiosperme viventi che hanno conservato caratteri di maggiore primitività appaiono difficilmente inquadrabili tra le dicotiledoni o le monocotiledoni. I dati più recenti sembrerebbero invece dimostrare che solo le monocotiledoni costituiscono un gruppo monofiletico (derivano cioè tutte da un unico antenato comune), mentre le dicotiledoni comprenderebbero almeno due gruppi a origine indipendente:
- il grande gruppo delle "vere dicotiledoni" (o eu-dicotiledoni) caratterizzate da avere polline con tre solchi (tricolpato), che comprende la maggior parte delle famiglie tradizionalmente inquadrate nelle dicotiledoni
- un gruppo molto più piccolo che include le famiglie che hanno mantenuto caratteri più primitivi. Questo gruppo, che corrisponde grosso modo alla sottoclasse Magnoliidae della classificazione di Cronquist, può essere ulteriormente suddiviso in due:
famiglie a portamento legnoso con foglie più o meno coriacee, come le Magnoliaceae (gruppo delle Magnoliidi arboree);
famiglie erbacee a foglie sottili, come le Nymphaeaceae e le Aristolochiaceae, con caratteristiche in parte simili a quelle delle monocotiledoni, di cui rappresenterebbero il gruppo ancestrale (Paleoerbe non monocotiledoni). Si tratta di un gruppo abbastanza eterogeneo.
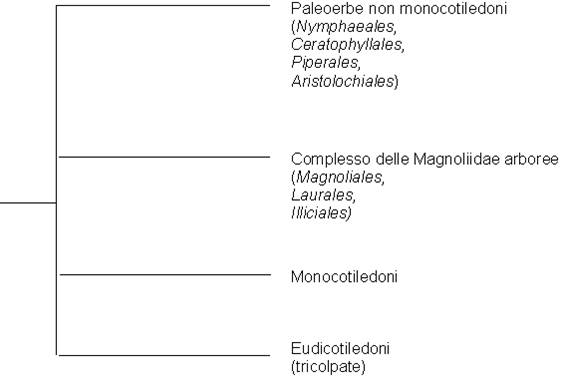
Fig. 6 - Classificazione delle angiosperme su base filogenetica (ridotto e modificato da Judd, Campbell, Kellogg, Stevens Botanica sistematica Ed. Piccin, 2002).
Tuttavia, per una sistematica elementare a carattere applicativo, la distinzione in monocotiledoni e dicotiledoni appare ancora utile e ad essa verrà fatto riferimento anche nella trattazione delle principali famiglie di interesse applicativo ed ecologico che si trova al termine del capitolo.
Seguendo il filo dell'evoluzione - 4: Da erba a albero e ritorno.
Le prime piante terrestri furono erbe. La pressione selettiva portò successivamente nelle pteridofite all'evoluzione della forma arborea (con relativo accrescimento secondario), che garantiva una migliore dispersione delle spore e consentiva di sfuggire all'ombreggiamento delle altre piante.
Anche le prime spermatofite o piante a seme furono alberi (pteridosperme) e arrivarono ad imporsi sulle contemporanee pteridofite arboree grazie alla maggiore efficienza dei processi riproduttivi legati alla comparsa dell'ovulo e del tubetto pollinico. Forme arboree di pteridofite (soprattutto licopodi) e in minor misura di gimnosperme dominarono la vegetazione del Carbonifero e della prima parte del Mesozoico (fino al Giurassico).
Nelle gimnosperme, che sono le spermatofite meno evolute, il legno secondario è di tipo omoxilo, formato cioè da elementi (tracheidi) a lume relativamente sottile che svolgono funzioni sia di conduzione che di sostegno. Questa doppia funzionalità delle tracheidi va però a scapito dell'efficienza nella conduzione e questo è uno dei motivi dell'aspetto più o meno xerofilo comune praticamente a tutte le gimnosperme. Molto più efficiente e versatile si rivelerà il legno eteroxilo delle angiosperme, dove i compiti di conduzione e di sostegno sono distribuiti tra i vari elementi costituenti (fibre, fibrotracheidi, tracheidi, trachee). Per la conduzione si dimostrano particolarmente efficaci le trachee, a lume ampio e prive di pareti trasversali. Bisogna tenere presente che il volume di liquido condotto aumenta con la quarta potenza del raggio del conduttore, il che significa che anche un piccolo aumento del lume cellulare degli elementi conduttori del legno porta ad un enorme aumento del volume di acqua trasportato: raddoppiando il raggio, il flusso si moltiplica per 16 volte; quadruplicando il raggio, per 256 volte. Questa efficienza di conduzione del legno eteroxilo rispetto al legno omoxilo è uno degli elementi che contribuirà all'affermazione delle angiosperme sulla terra.
Probabilmente, anche le prime angiosperme furono arboree, anche se secondo alcuni paleobotanici la maggior parte delle linee filetiche delle antiche angiosperme comprendono sia specie erbacee che legnose. Con una certa approssimazione si può dire che fino alla loro comparsa il cammino dell'evoluzione delle piante terrestri sia andato costantemente nella direzione di una sempre maggiore altezza e longevità dei singoli individui. È nell'ambito delle angiosperme che la tendenza cambia e si assiste ad un ritorno secondario verso l'abito erbaceo e addirittura verso le piante a ciclo di vita annuale.
Piccolo è bello: i vantaggi dell'abito erbaceo e del ciclo vitale breve.
Che una pianta alta e longeva possa avere dei vantaggi evolutivi rispetto a una pianta erbacea è intuitivo: ogni anno vincendo la concorrenza delle piante più basse manda più lontano i suoi elementi di propagazione (spore nelle pteridofite, semi nelle spermatofite) a colonizzare nuovi spazi; inoltre riesce a intercettare la luce a discapito di chi rimane negli strati inferiori ombreggiati.
Più difficile è capire perché a un certo punto si sia rivelato vantaggioso il ritorno alle piccole dimensioni legate a un ciclo di vita breve. Eppure bisogna considerare che:
- un albero impiega anni a raggiungere l'età riproduttiva. In tutto questo tempo non produce nuovi individui, cioè nuove combinazioni geniche che possano essere sperimentati dall'evoluzione. E ci sono molte possibilità che muoia prima di arrivare all'età riproduttiva e alle dimensioni definitive. Una pianta erbacea invece comincia a produrre semi fin dal primo anno, o dopo soli pochi anni di vita vegetativa.
- anche quando è adulto e produce semi ogni anno, un albero dà una progenie che deriva sempre dallo stesso materiale genetico di partenza (lo sporofito genitore), quindi con ridotte possibilità di variazioni rispetto a questo e agli individui "fratelli". In una pianta erbacea a ciclo breve, dopo qualche anno di produzione di semi (un solo anno nel caso estremo delle annuali), la pianta muore e viene sostituita da discendenti che hanno patrimonio genetico variato. Le generazioni si susseguono quindi molto più velocemente, con una possibilità enormemente più grande di comparsa di nuovi caratteri e in definitiva di evoluzione di nuove forme che si possono rivelare più adatte all'ambiente o a colonizzarne di nuovi.
Se dunque la longevità è un vantaggio per il singolo individuo, non lo è ai fini dell'evoluzione della specie (o più in generale del gruppo) di cui quell'individuo fa parte. Ecco un altro vantaggio evolutivo delle angiosperme: nei gruppi che comprendono rappresentanti a ciclo vitale breve, si sono potute evolvere forme e funzioni adatte agli ambienti più diversi in un tempo relativamente rapido. Basta considerare come appare limitata la varietà morfologica e ambientale presente in una famiglia di gimnosperme arboree come le Pinaceae se la si confronta con quella di angiosperme erbacee come le Poaceae (o Gramineae), diffuse a tutte le latitudini e nei più diversi ambienti: praterie, steppe, savane, sottobosco di foreste dei più diversi tipi, ambienti antropizzati, ecc. Nessuna famiglia di gimnosperme presenta un grado di diversità paragonabile.
Sui testi:
RAVEN, EVERT, EICHHORN
Cap. 21 Introduzione alle angiosperme
(p. 509-528)
Cap. 22 Evoluzione delle angiosperme
(p. 529-565)
Cap. 23 Prime fasi di sviluppo della pianta
(p. 570-583)
Cap. 25 La radice
generalità, gli apparati radicali (p. 603-607)
Cap. 26 Il germoglio
Le foglie sono disposte Morfologia della foglia (p. 641-642)
Modificazioni del fusto (p. 658-661)
CRONQUIST:
Cap. 23 Introduzione alle Angiosperme
(p. 349-350)
Cap. 24 Angiosperme: fusti
Evoluzione dei tipi di fusti, Fusti modificati, Riproduzione indotta (p. 375-381)
Cap. 25 Angiosperme: radici
Posizione, funzioni, tipi (p. 383-385)
Radici modificate (p. 394-399)
Cap. 26 Angiosperme: foglie
Struttura esterna (p. 401-407)
Foglie specializzate (p. 412-414)
Cap. 27 Angiosperme: rapporti con l'acqua
Idrofite, mesofite, xerofite (p. 427-430)
Cap. 31 Angiosperme: fiori
Introduzione, differenziazione (p. 484)
Struttura dei fiori: organi, tipi di fiori, infiorescenze (p. 488-490, 491-494)
Riproduzione (p. 494-503)
Cap. 32 Angiosperme: frutti, semi e plantule
Frutti, dispersione semi, struttura seme (p. 507-513)
Cap. 34 Angiosperme: classificazione
Introduzione, Principi, Dicotiledoni e Monocotil., Cenni storici, Tendenze evolutive (p. 546-555)
Cap. 35 Angiosperme: importanza economica
(E' consigliata la lettura del capitolo)
Cap. 38 L'evoluzione
Storia, Alcuni concetti, Differenziazione: ruolo dell'isolamento (p. 622-627)
Vita primitiva, Alghe (introduzione), Funghi, Embriofite (p. 629-630, 632-638)
Seguendo il filo dell'evoluzione - 5: La riproduzione sessuale nelle piante terrestri
Tutte le piante terrestri (embriofite) sono oogame. L'oosfera si sviluppa all'interno di un gametangio femminile (archegonio) circondato da uno strato di cellule sterili, e non semplicemente avvolto dalla parete cellulare come nelle alghe.
Nel corso dell'evoluzione delle piante terrestri, l'archegonio va incontro ad un processo di progressiva riduzione, fino a scomparire nelle angiosperme, dove le due cellule sinergidi ne rappresentano a parere di molti le ultime vestigia. In tutte le embriofite, nelle prime fasi di accrescimento dopo la formazione dello zigote (embrione), lo sporofito è parassita del gametofito.
Questa dipendenza è completa e dura praticamente per l'intera durata di vita dello sporofito nelle briofite, dove lo sporofito svolge una limitatissima attività fotosintetica e affonda la sua parte basale nei tessuti del gametofito, da cui trae nutrimento. Solo in alcune briofite molto specializzate (Anthoceros) lo sporofito ha durata di vita pluriennale, arriva a contatto diretto con il terreno per mezzo di un piede e sembra assumere una certa capacità di vita autotrofa autonoma.
Nei gruppi di piante più evoluti, lo sporofito è la generazione dominante, diventa perenne e la sua dipendenza trofica dal gametofito diventa temporanea, dal momento che si affranca dal gametofito in una fase sempre più precoce del suo sviluppo. Parallelamente, si assiste alla riduzione dell'entità e della durata di vita del gametofito.
Nelle pteridofite, il gametofito (protallo) è ancora capace di vita autonoma e nutre il nuovo sporofito fino a che questo non si rende autonomo sia per l'assorbimento dell'acqua dal terreno che per la fotosintesi. A questo punto, il gametofito ha esaurito il suo compito e di regola muore. Eppure, nonostante il ribaltamento dei ruoli fra le due generazioni rispetto alle briofite e il miglior adattamento dello sporofito alla vita in ambiente terrestre grazie alla comparsa dei tessuti vascolari, le pteridofite non riescono ad emanciparsi del tutto dall'acqua e la loro diffusione rimane confinata ad ambienti umidi. Questo a causa di alcuni aspetti del loro ciclo riproduttivo che condividono con le briofite:
- l'acqua - anche se in piccola quantità - è ancora necessaria al momento della gamia, perché consente ai gameti maschili flagellati di nuotare per breve tratto fino agli archegoni, dove si trovano i gameti femminili immobili.
- l'organo di diffusione è ancora la meiospora, obbligata a germinare dove esistono le condizioni di umidità necessarie per la crescita del gametofito. Lo sporofito nasce nell'archegonio e per tutta la durata della sua vita sarà condannato a vegetare là dove si era sviluppato il gametofito. Pur essendo la generazione meno sviluppata e dalla vita più breve, è il gametofito che "sceglie" dove crescerà la nuova pianta, e non lo sporofito, che pure dispone di adattamenti che consentono la vita anche al di fuori dell'ambiente umido (radici, tessuti di conduzione).
Entrambi questi limiti vengono definitivamente superati nelle spermatofite (piante a seme), le prime in grado di colonizzare anche ambienti aridi:
- con la comparsa dell'ovulo e del granulo polinico, l'oosfera raggiunge la massima protezione e nessuno dei due gameti viene più liberato all'esterno. Il gametofito maschile viene trasportato in vicinanza di quello femminile dal granulo pollinico e attraverso il tubetto il gamete maschile può raggiungere e fecondare quello femminile senza mai essere esposto all'aria. L'ambiente umido necessario all'incontro dei gameti viene ricreato all'interno dei tessuti della pianta. Il mare in cui nuotano e si incontrano i gameti degli organismi acquatici si riduce nelle piante terrestri dapprima a un velo di rugiada (briofite, pteridofite), poi alla microscopica quantità di liquido contenuto nella camera archegoniale (Cycadopsida, Gynkgoopsida), per scomparire definitivamente, insieme ai flagelli dei gameti maschili, nelle gimnosperme più evolute e nelle angiosperme.
- l'organo di diffusione è il seme, giovane sporofito quiescente fornito di tessuti di riserva e protetto da tegumenti. Sarà questo a "scegliere" il luogo e l'ambiente dove crescerà la nuova pianta, al di fuori di ogni condizionamento da parte del gametofito. Inoltre il seme, organo pluricellulare con tessuti differenziati, potrà sviluppare strutture che facilitino e rendano più efficiente la diffusione (ali, ecc.).
Nelle angiosperme, i processi riproduttivi raggiungono il massimo livello di perfezionamento e di efficienza nel mondo vegetale, grazie a due fondamentali innovazioni premiate dalla selezione: il carpello e il fiore.
Il carpello presenta una serie di notevoli vantaggi evolutivi:
- ulteriore protezione del gametofito femminile dal pericolo di disseccamento e di danneggiamenti;
- possibilità di aumentare il periodo di ricettività del polline, senza rischio di disseccamento per l'ovulo;
- grazie all'allontanamento della superficie recettiva dall'oosfera, possibilità per la pianta di mettere in atto meccanismi di incompatibilità sul percorso più lungo del tubetto pollinico, arrivando in definitiva a scegliere il gamete maschile più "gradito" per la fecondazione;
- possibilità di contribuire attivamente alla dispersione del seme, con la trasformazione del carpello in frutto dopo la fecondazione e le relative modificazioni in funzione dell'agente della dispersione (frutti carnosi ingeriti dagli animali, frutti alati portati dal vento, ecc.).
Nel fiore gli organi e le strutture riproduttive sono organizzati in un complesso estremamente specializzato che garantisce la massima protezione ai gametofiti e la massima efficienza nella selezione e nell'incontro dei gameti.
Nelle piante a fiore raggiunge dunque la massima espressione quella tendenza evolutiva verso il perfezionamento dei meccanismi riproduttivi che è alla base della possibilità di insorgenza di nuove forme e nuove funzioni da sottoporre al vaglio della selezione. È proprio grazie a questo che tra le angiosperme si è potuta evolvere quell'enorme varietà morfologica e fisiologica che ha loro consentito di diffondersi e diventare il gruppo di vegetali attualmente dominante in quasi tutti gli ambienti terrestri.
L'anatomia e la fisiologia delle piante a fiore sono affrontati nel corso di Biologia generale. Le caratteristiche morfologiche degli organi vegetativi (radici, fusti, foglie) e riproduttivi (fiori, frutti) vengono considerate nelle esercitazioni del modulo di Botanica sistematica e per il loro approfondimento si rimanda alla relativa Guida alla descrizione. Le caratteristiche vegetative e l'interesse economico ed ecologico delle angiosperme arboree di interesse forestale vengono trattate nel modulo di Botanica forestale.
|
| Appunti su: magnoliophyta ciclo biologico, |
|