|
| Appunti scientifiche |
|
|
| Appunti scientifiche |
|
| Visite: 7475 | Gradito: |
Leggi anche appunti:La chimica e il metodo scientifico d'indagineLA CHIMICA E IL METODO SCIENTIFICO D'INDAGINE Noi, nella catena alimentare, La legge di proustLA LEGGE DI PROUST OBIETTIVO: verificare che gli elementi L'ACQUA - Struttura chimicaL'ACQUA Struttura chimica L'acqua è una delle sostanza più importanti |
 |
 |
I primi esperimenti sul meccanismo dell'eredità furono compiuti verso il 1860 da Gregor Mendel (1882-1884), in un monastero nell'attuale Brno, eseguendo incroci su piante di pisello (Pisum sativum). I risultati ottenuti da Mendel vengono tradizionalmente riassunti in alcune leggi (le leggi di Mendel). Se si confrontano i diversi manuali di genetica si noterà che la formulazione delle leggi varia sia per quanto riguarda il numero (due o tre leggi) che per ciò che riguarda l'ordine di enunciazione e la loro denominazione. In effetti Mendel non formulò alcuna legge, ma i suoi risultati sono più agevolmente esprimibili e riassumibili sotto forma di enunciati.
Inoltre, nell'enunciare queste due o tre leggi, la maggior parte dei trattati di genetica è costretta a far uso dei concetti di gene, allele, omozigote ed eterozigote. Si tenga comunque presente che tali concetti erano totalmente estranei a Mendel. Infatti i termini allelomorfo, omozigote ed eterozigote furono introdotti solo nel 1902 da Bateson, mentre il termine gene fu coniato da Johannsen nel 1909.
Mendel si occupò di caratteri che si presentavano nel fenotipo secondo modalità ben distinte che non ammettevano forme intermedie. I risultati ottenuti da Mendel si applicano dunque solo a questo tipo di caratteri, noti come caratteri mendeliani o caratteri discreti o caratteri a variabilità discontinua. Si tenga presente che tali caratteri non sono molto diffusi in natura. La maggior parte dei caratteri sono infatti caratteri a variabilità continua, come l'altezza di un individuo, il colore della pelle etc.
La pianta di pisello (Pisum sativum), su cui lavorò Mendel, risultò particolarmente adatta per questo tipo di esperimenti. È infatti di facile coltivazione, può dare più di una generazione all'anno, presenta parecchi caratteri a variabilità discontinua, ma soprattutto si riproduce per autofecondazione. Il fiore infatti non si schiude finché i gameti maschili (polline) non abbiano fecondato gli ovuli femminili dello stesso fiore (ricordiamo che anche la pianta di pisello, come la maggior parte dei vegetali, è ermafrodita, presenta cioè sia l'apparato riproduttore maschile che quello femminile riuniti in una medesimo individuo).
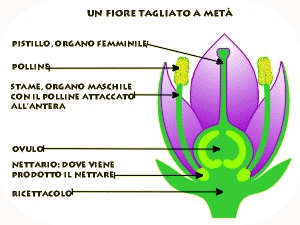
Non potendo dunque ricevere polline da altre piante (fecondazione incrociata), la pianta di pisello non riceverà neppure caratteri diversi da quelli propri. In queste condizioni si osserva che i discendenti di un certo individuo presenteranno sempre i medesimi caratteri, identici a quelli dei genitori. Gli individui di questo tipo costituiscono una linea pura. Per Mendel fu dunque relativamente semplice isolare linee pure che differivano per un carattere ed incrociarle, praticando su di esse la fecondazione artificiale. L'operazione richiede che il fiore venga aperto e ne vengano recisi gli stami contenenti il polline per evitare l'autofecondazione. Successivamente l'organo femminile viene fecondato con il polline proveniente da un'altra pianta. Se si incrociano due linee pure che differiscono per un carattere gli individui che si ottengono non appartengono ovviamente più ad una linea pura e vengono definiti ibridi (monoibridi).
Le ricerche sui piselli impegnarono MENDEL dal 1854 al 1864. In questo periodo egli coltivò e analizzò almeno 28.000 piante della specie Pisum Sativum, prendendo in considerazione sette coppie di caratteri del seme e della pianta quali l' altezza, la posizione dei fiori, la forma e il colore dei semi.
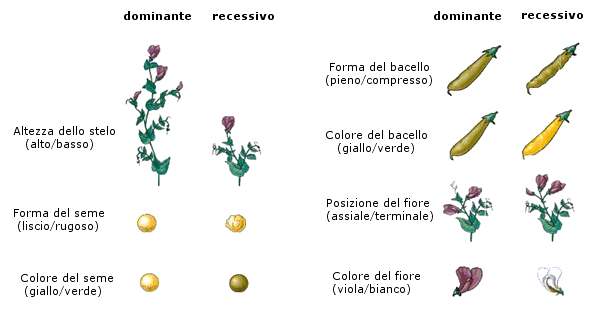
Mendel incrociò fra loro a due a due linee pure che differivano per un carattere (pianta alta x pianta bassa, pianta con semi verdi x pianta con semi gialli, etc.), classificando e contando gli esemplari ottenuti in ciascuna generazione e cercando di individuare l' esistenza di precisi rapporti matematici tra i diversi individui presenti ad ogni generazione.
I risultati di questi incroci portarono Mendel a conclusioni in netto contrasto con il modello generalmente accettato allora dell' eredità per rimescolamento, secondo il quale "essenze" di ciascuno dei due genitori, trasmesse attraverso liquidi simili al sangue, contribuivano "mescolandosi" alla formazione dei figli. Mendel avanzò invece l'ipotesi che l' eredità fosse "particolata", cioè che i diversi caratteri ereditari fossero portati da strutture biologiche discrete e distinte, unità di eredità (o fattori unitari), anche se a quel tempo non vi erano prove dirette per dimostrarne l' esistenza, e che noi oggi chiamiamo geni ed alleli.
Egli aveva infatti notato che, incrociando piante di piselli che si differenziavano per singoli caratteri facilmente individuabili, non si ottenevano ibridi con caratteri mescolati, ma la generazione F1 era formata da individui che presentavano sempre il carattere di uno solo dei due genitori. In termini moderni diremo che il carattere che si manifesta è dominante, quello che rimane nascosto è recessivo.
In uno dei suoi esperimenti, Mendel incrociò piante di pisello che da molte generazioni producevano solo fiori violetti con piante che da molte generazioni producevano solo fiori bianchi. Egli usò il polline prodotto da piante a fiori violetti per fecondare piante a fiori bianchi. Fece anche l'operazione inversa, cioè fecondò con polline proveniente da piante a fiori bianchi piante a fiori violetti. Il risultato era sempre il medesimo: in F1 gli ibridi presentavano sempre fiori violetti.
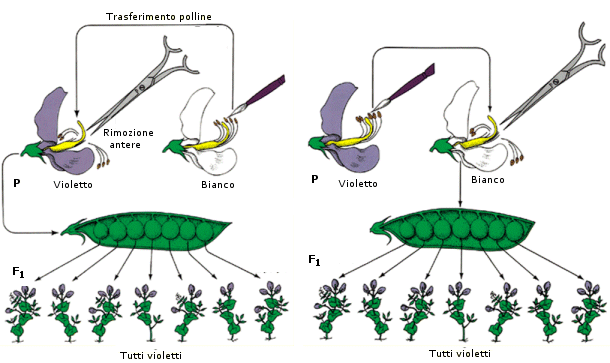
I risultati di questi incroci possono essere riassunti in termini moderni nella legge della dominanza o legge dell'uniformità degli ibridi F1: l'incrocio tra un omozigote dominante (linea pura dominante) ed un omozigote recessivo (linea pura recessiva) genera una generazione F1 di eterozigoti (ibridi) fenotipicamente uguali al genitore dominante.
P Linea pura dominante x Linea pura recessiva
F1 Ibrido dominante
Si tenga presente che una linea pura, presentando un solo tipo di carattere, corrisponde ad un omozigote, mentre un ibrido, in cui si sono uniti i caratteri provenienti da due linee pure, corrisponde ad un eterozigote.
In una successiva serie di esperimenti, Mendel incrociò tra loro gli ibridi F1. In realtà l'incrocio tra gli ibridi F1 non richiede una fecondazione artificiale. Infatti, lasciando che gli ibridi si autofecondino si ottiene un incrocio tra ibridi.
La progenie, seconda generazione filiale (F2) risultò composta per il 75% di individui fenotipicamente dominanti e per il 25% di individui fenotipicamente recessivi (rapporto 3:1). Ricompare il carattere recessivo presente nell'incrocio parentale e scomparso in F1.
Per spiegare i risultati ottenuti Mendel avanzò alcune ipotesi, poi rivelatesi corrette:
Oggi noi sappiamo che il concetto di 'segregazione' dei due fattori genetici e successiva ricongiunzione casuale ipotizzato da Mendel coincide con i processi meiotici di gametogenesi (in cui si separano gli omologhi e quindi gli alleli da essi portati) e di fecondazione (in cui i gameti si combinano casualmente a formare lo zigote).
La riproduzione sessuale avviene infatti grazie alla formazione di gameti (ovuli e spermatozoi), cellule aploidi che si formano tramite meiosi, un processo in cui i cromosomi omologhi, che ospitano i due alleli di un carattere, si separano. Successivamente, durante la fecondazione, l'unione casuale dei gameti produce la ricongiunzione degli alleli.
Quando si osservano i risultati di un incrocio, si misurano le frequenze con cui compaiono determinati caratteri. Se, ripetendo più volte il medesimo incrocio, le frequenze con cui compaiono certi caratteri si attestano sempre intorno a determinati valori, allora possiamo utilizzare questi valori come una stima della probabilità che quel determinato risultato si verifichi come conseguenza dell'incrocio in questione.
Un esempio servirà a chiarire.
Se lancio 100 volte una moneta in aria potrò ottenere 55 volte 'testa' e 45 volte 'croce'. 'Testa' compare con una frequenza del 55%. 'Croce' compare con una frequenza del 45%.
Se rifaccio più volte i 100 lanci mi accorgerò che le frequenze con cui compaiono 'testa' e 'croce' oscillano sempre intorno al 50%, anche se difficilmente saranno esattamente pari al 50%. Posso anche facilmente verificare che le frequenze con cui compaiono i due eventi (Testa e Croce) si avvicineranno tanto più al 50% quanto più alto sarà il numero di lanci.
In effetti la frequenza è una grandezza che si misura ed è il rapporto tra il numero di volte che un determinato evento effettivamente si verifica ed il numero totale di prove effettuate (Frequenza = rapporto tra gli eventi favorevoli osservati e gli eventi totali)
La probabilità è invece una grandezza che si calcola ed è il rapporto tra il numero di eventi favorevoli ed il numero di eventi possibili.
La probabilità che quando lancio una moneta esca 'testa' è del 50% (0,5) poiché vi è solo un caso favorevole (solo 1 lato della moneta presenta 'testa') mentre 2 sono i casi possibili (due sono le facce della moneta). Ma anche se la probabilità che esca testa è pari allo 0,5 (50%), la frequenza con cui uscirà 'testa' se lancio effettivamente la moneta difficilmente sarà sempre del 50%.
Sperimentalmente si osserva tuttavia che la frequenza di un evento si avvicina tanto più al valore calcolato della sua probabilità quanto più elevato è il numero di eventi osservati (è più probabile che la frequenza con cui esce 'testa' si avvicini al 50% se effettuo 1000 lanci piuttosto che se ne effettuo 10).
Da questa ultima osservazione scaturisce un'altra definizione di probabilità: la probabilità di un evento è il valore al quale tende la frequenza quando il numero degli eventi osservati tende ad infinito.
Questo in definitiva è anche il motivo per il quale, per stimare la probabilità che da un incrocio si generi un particolare carattere piuttosto che un altro, dobbiamo incrociare un numero molto elevato di individui.
È allora ovvio che quando Mendel lasciò che i monoibridi F1 si incrociassero non ottenne mai esattamente il 75% di individui dominanti ed il 25% di individui recessivi, ma misurò frequenze che oscillavano intorno a questi valori e ne desunse che le probabilità dovevano essere del 75% e del 25%.
È tuttavia possibile, dato un certo incrocio, calcolare le probabilità dei genotipi risultanti senza dover materialmente effettuare l'incrocio.
Per farlo dobbiamo conoscere i genotipi degli individui che si incrociano e simulare gli eventi che portano alla formazione dei nuovi individui: gametogenesi e fecondazione.
Si calcolano quindi le probabilità che durante la gametogenesi gli alleli del carattere in questione si trovino su di un determinato gamete e successivamente si calcolano le probabilità, attraverso un quadrato di Punnett, che i gameti femminili si 'incontrino' con i gameti maschili.
Il quadrato di Punnett è una tabella a doppia entrata che serve a simulare tutte le possibili combinazioni dei gameti maschili con quelli femminili (fecondazione) e a calcolare la probabilità dei genotipi che si generano come conseguenza dell'unione dei gameti. I gameti maschili (con le loro probabilità) vengono posti ad un lato del quadrato ed i gameti femminili (con le loro probabilità) vengono posti su di un lato adiacente al precedente. All'interno del quadrato vengono poi effettuate (come nel gioco della battaglia navale) tutte le combinazioni tra le righe e le colonne. Le probabilità dei genotipi che si realizzano non sono altro che la probabilità di un evento composto, cioè la probabilità che due eventi semplici (il gamete maschile e quello femminile) si verifichino contemporaneamente.
La probabilità di un evento composto si calcola come il prodotto delle probabilità degli eventi semplici che concorrono alla sua uscita. Così le probabilità dei genotipi che si formano all'interno del quadrato di Punnett si calcola come prodotto delle probabilità dei singoli gameti che devono combinarsi per dare quel determinato genotipo.
Ad esempio la probabilità che lanciando contemporaneamente un dado ed una moneta esca il numero 5 sul dado e 'testa' sulla moneta è pari a 1/6 x 1/2 = 1/12. La probabilità che esca il 6 è infatti 1/6, la probabilità che esca 'testa' è 1/2, la probabilità che escano contemporaneamente (evento composto) è 1/12.
Proviamo allora a simulare l'incrocio tra due ibridi F1 a fiori violetti. Si tratta di due eterozigoti con genotipo Pp, (con P = purple, violetto = allele dominante e p = bianco = allele recessivo)
Per prima cosa dobbiamo simulare la gametogenesi (meiosi). Tenendo presente che l'allele dominante P si trova su di un cromosoma e l'allele recessivo p sul suo omologo e che durante la meiosi gli omologhi si separano ordinatamente a metà, avremo metà gameti che contengono l'allele dominanti e metà l'allele recessivo. In termini di probabilità potremo dire che ogni individuo eterozigote che partecipa all'incrocio ha una probabilità del 50% (0,5) di produrre gameti con l'allele dominante e del 50% (0,5) di produrre gameti con l'allele recessivo (si ricordi che la probabilità è teorica. Le frequenze con cui si generano realmente i gameti possono essere diverse, ad esempio per problemi di sopravvivenza degli stessi).
Una volta creati i gameti, li sistemiamo ai due lati del quadrato di Punnett con le loro probabilità. Da un lato i gameti maschili (♂) e dall'altro i gameti femminili (♀).
Ora costruiamo all'interno del quadrato tutte le 4 possibili combinazioni di gameti maschili e femminili. Questi sono i 4 genotipi che possono prodursi dall'incrocio. Ognuno di essi avrà una probabilità di formarsi del 25% (0,5 x 0,5 = 0,25)

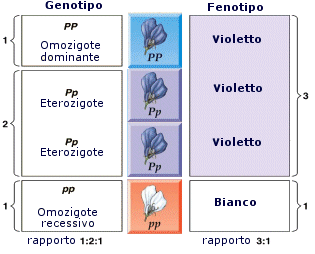
Dall'incrocio di due ibridi si possono formare dunque un 25% di omozigoti dominanti (PP), un 50% di eterozigoti (Pp) ed un 25 % di omozigoti recessivi (pp). Si noti come, essendo gli omozigoti dominanti e gli eterozigoti fenotipicamente uguali, l'incrocio produce un 75% di individui fenotipicamente dominanti ed un 25% di individui recessivi. Lo stesso risultato ottenuto sperimentalmente da Mendel.
I risultati di questa seconda serie di incroci possono dunque essere riassunti in termini moderni nella legge della segregazione o legge della disgiunzione: l'incrocio tra due eterozigoti (ibridi) genera una generazione F2 costituita da un 25% di omozigoti recessivi, da un 50% di eterozigoti e da un 25% di omozigoti dominanti. Il rapporto fenotipico dominanti/recessivi è 3:1
F1 Ibrido x Ibrido
F2 75% dominanti 25% recessivi
Come abbiamo già anticipato, le leggi di Mendel non sono generali, ma presentano delle eccezioni (dominanza incompleta e codominanza) Non sempre esistono infatti un allele dominante ed uno recessivo per un dato carattere.
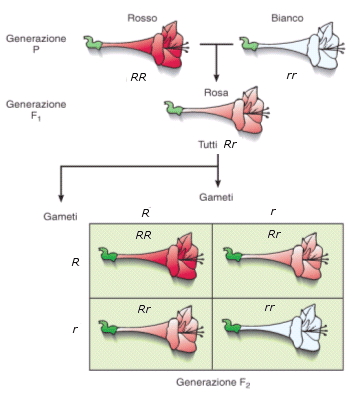 La pianta della bella di notte (Mirabilis jalapa) può avere
fiori rossi, bianchi o rosa. Le piante con fiori rossi hanno due copie
dell'allele R per il colore rosso dei fiori e sono, quindi, omozigoti dominanti
RR. Le piante con fiori bianchi hanno due copie dell'allele r per
il colore bianco dei fiori e sono, quindi, omozigoti recessive rr. Le
piante con una copia di ciascun allele, cioè gli eterozigoti Rr, hanno
fenotipo rosa, intermedio tra i colori prodotti dai due alleli. Questo è un
esempio del fenomeno di dominanza incompleta.
La pianta della bella di notte (Mirabilis jalapa) può avere
fiori rossi, bianchi o rosa. Le piante con fiori rossi hanno due copie
dell'allele R per il colore rosso dei fiori e sono, quindi, omozigoti dominanti
RR. Le piante con fiori bianchi hanno due copie dell'allele r per
il colore bianco dei fiori e sono, quindi, omozigoti recessive rr. Le
piante con una copia di ciascun allele, cioè gli eterozigoti Rr, hanno
fenotipo rosa, intermedio tra i colori prodotti dai due alleli. Questo è un
esempio del fenomeno di dominanza incompleta.
Reincrociando tra loro gli individui F1, la F2 presenterà tre classi fenotipiche in rapporto 1:2:1 (1/4 a fiore rosso, 2/4 a fiore rosa e 1/4 a fiore bianco) anziché di 3:1.
Il reincrocio è una pratica della genetica sperimentale usata per determinare il genotipo incognito di una popolazione fenotipicamente dominante, la quale potrebbe essere composta sia da omozigoti dominanti che da eterozigoti.
La popolazione dominante viene incrociata con individui omozigoti recessivi (fenotipo recessivo). Si può dimostrare che la percentuale di eterozigoti presenti nella popolazione da testare è sempre il doppio della percentuale di recessivi che si ottengono dal test-cross.
In particolare la percentuale di recessivi che si ottengono dal reincrocio può andare dallo 0% (nessun eterozigote: la popolazione da testare è tutta omozigote dominante) al 50% (la popolazione da testare è tutta eterozigote: il doppio di 50%).
Si prenda ad esempio una popolazione di cavie a pelo nero formata da una frazione X di omozigoti dominanti per il colore del pelo (BB dove B = Black, nero) ed una frazione Y di eterozigoti (Bb) (ovviamente X+Y = 1). Si incrocino ora gli individui con una popolazione di cavie recessiva a pelo marrone formata da omozigoti recessivi (bb)
Simuliamo ora la gametogenesi e calcoliamo le probabilità di formazione dei diversi gameti.
Gli omozigoti dominanti produrranno solo gameti contenenti l'allele B e quindi formeranno una quantità X di gameti B
Gli eterozigoti produrranno metà gameti B e metà gameti b e quindi formeranno una quantità Y/2 di gameti B ed Y/2 di gameti b
Gli omozigoti recessivi produrranno un 100% (e quindi una frazione pari ad 1) di gameti b.
Inserendo ora questi dati nel quadrato di Punnett si osserva che si genera una frazione Y/2 di omozigoti recessivi (bb), esattamente la metà degli Y eterozigoti presenti nella popolazione da testare

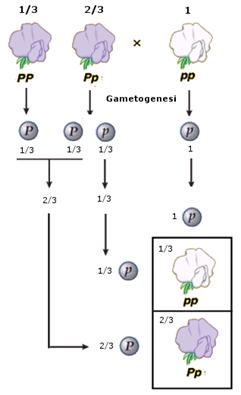
Il reincrocio fu utilizzato da Mendel per determinare la percentuale di omozigoti dominanti e di eterozigoti che costituivano il 75% di individui fenotipicamente dominanti che si presentavano in F2 dall'incrocio di due ibridi.
Se, ad esempio, effettuiamo un reincrocio del 75% di individui dominanti a fiori violetti (una popolazione che sappiamo formata per 2/3 (il 50% di 75%) da eterozigoti Pp ed 1/3 (il 25% di 75%) da omozigoti dominanti PP) con una popolazione recessiva a fiori bianchi, otteniamo tipicamente 1/3 di individui a fiori bianchi (la metà degli eterozigoti presenti nella popolazione da testare).
Quando ebbe chiarito come avveniva la trasmissione di un singolo carattere, Mendel studiò la trasmissione di due caratteri alla volta, sempre effettuando incroci controllati e registrando tutti i risultati ottenuti.
Mendel eseguì un incrocio tra piante di linee pure che differivano contemporaneamente per due caratteri, per esempio forma e colore del seme (una linea pura a seme giallo e liscio ed una linea pura a seme verde e rugoso).
Si trattava di capire se i due caratteri venivano sempre ereditati insieme o se potevano essere trasmessi indipendentemente uno dall'altro. In altre parole, una pianta che presenta i semi gialli dovrà necessariamente ereditare anche il carattere 'seme liscio' o potrà presentare anche il carattere 'seme rugoso'?
La generazione F1 risultò costituita esclusivamente da individui ibridi (diibridi) sia per la forma che per il colore del seme, con semi gialli e lisci. Questo risultato permetteva solo di affermare che la forma liscia è dominante sulla forma rugosa così come il colore giallo è dominante sul colore verde.
Effettuata l'autofecondazione di questi ibridi Mendel ottenne una generazione F2 in cui erano presenti quattro tipi di semi nel rapporto di 9:3:3:1 e precisamente: semi gialli e lisci (9), semi verdi e lisci (3), semi gialli e rugosi (3), semi verdi e rugosi (1).
La comparsa di combinazioni fenotipiche nuove (ricombinanti), non presenti nella generazione parentale (verdi-lisci e gialli-rugosi), con caratteri scambiati, permise a Mendel di affermare che le coppie di caratteri sono indipendenti e la trasmissione ad un discendente di uno o l'altro degli alleli di un carattere non influenza la probabilità di trasmettergli uno o l'altro degli alleli per l'altro carattere.
I risultati di questi incroci vengono solitamente sintetizzati nella legge dell'indipendenza dei caratteri o legge dell'assortimento indipendente: L'incrocio tra due linee pure che differiscono per due caratteri produce una generazione F1 di ibridi per entrambi i caratteri che, incrociati danno una generazione F2 in cui sono presenti tutte le combinazioni fenotipiche nella proporzione 9 D/D : 3 D/R : 3 R/D : 1 R/R (D = fenotipo dominante; R = fenotipo recessivo).
In realtà Mendel aveva studiato un caso particolare. Le coppie di caratteri analizzate da Mendel si trovavano infatti su diverse coppie di omologhi e questo permettavo loro, durante la meiosi di segregare l'una indipendentemente dall'altra (assortimento indipendente).
Se una coppia di caratteri è portata dalla medesima coppia di omologhi, i due alleli appartenenti a caratteri diversi, che si trovano sul medesimo omologo tendono ovviamente ad essere trasmessi insieme. In questo caso i caratteri si dicono associati o linked e, come vedremo, non forniscono in F2 la medesima distribuzione di frequenza (9:3:3:1) travata da Mendel.
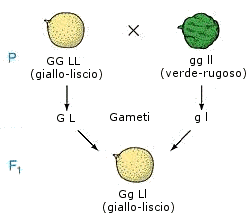 L'incrocio parentale produrrebbe ibridi eterozigoti per entrambi i caratteri
sia nel caso di caratteri indipendenti (portati da coppie diverse di omologhi)
che per caratteri associati (portati dalla medesima coppia di omologhi) è
infatti evidente che l'individuo GG LL (omozigote dominante per entrambi i
caratteri) non può produrre che gameti G
L, mentre l'individuo gg ll (omozigote recessivo per entrambi i caratteri)
non può produrre che gameti g l.
Dall'unione di questi due gameti si produrranno necessariamente un 100% di
eterozigoti per entrambi i caratteri Gg Ll.
L'incrocio parentale produrrebbe ibridi eterozigoti per entrambi i caratteri
sia nel caso di caratteri indipendenti (portati da coppie diverse di omologhi)
che per caratteri associati (portati dalla medesima coppia di omologhi) è
infatti evidente che l'individuo GG LL (omozigote dominante per entrambi i
caratteri) non può produrre che gameti G
L, mentre l'individuo gg ll (omozigote recessivo per entrambi i caratteri)
non può produrre che gameti g l.
Dall'unione di questi due gameti si produrranno necessariamente un 100% di
eterozigoti per entrambi i caratteri Gg Ll.
L'incrocio dei diibridi F1 tramite autofecondazione è invece più complesso. Infatti, trattandosi di caratteri indipendenti, il carattere colore del seme è portato da una coppia di omologhi, mentre il carattere forma del seme è portato da un'altra coppia di omologhi. Durante la gametogenesi Il cromosoma che contiene l'allele G ha il 50% di probabilità di segregare in un gamete con il cromosoma che contiene l'allele L e il 50% di probabilità di segregare con il cromosoma che contiene l'allele l. Ovviamente questo vale anche per l'altro omologo, che contiene l'allele g. In questo modo, l'assortimento indipendente degli omologhi genera 4 tipi di gameti: GL, Gl, gL, gl, ciascuno con una probabilità del 25% (1/4).
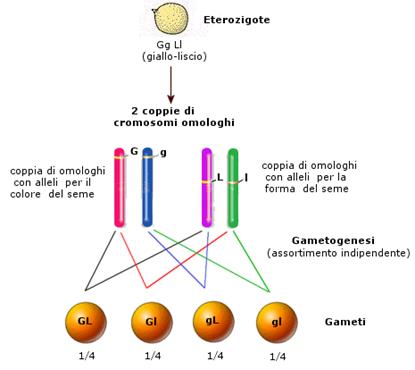
Se ora sistemiamo questi 4 tipi di gameti in un quadrato di Punnett, possiamo ottenere tutti i genotipi possibili.

In effetti i due caratteri vengono trasmessi come fossero due cararatteri singoli e obbediscono pertanto alla seconda legge di Mendel (legge della segregazione). Per ognuno di essi ritroviamo infatti una distribuzione 3:1 (75% di dominanti e 25% di recessivi) tipica dell'incrocio di un monoibridi. La distribuzione 9:3:3:1 di un di ibrido nasconde due distribuzioni 3:1 di due monoibridi.
Infatti se prendiamo in considerazione il primo carattere (colore del seme), su 16 individui, 12 sono dominanti (gialli) e 4 sono recessivi (verdi), con un rapporto fenotipico di 12:4 = 3:1.
Se invece prendiamo in considerazione il secondo carattere (forma del seme), su 16 individui, 12 sono dominanti (lisci) e 4 sono recessivi (rugosi), con un rapporto fenotipico di 12:4 = 3:1

Vediamo ora cosa ci saremmo dovuti attendere se i due caratteri fossero stati associati, se fossero stati cioè ospitati sulla medesima coppia di omologhi. In questo caso, per un generico diibrido AaBb, si sarebbero potuti formare solo due tipi di gameti (e non 4 come nel caso di caratteri indipendenti). I due caratteri si sarebbero comportati come un unico carattere, generando una distribuizione 3:1, con il 75% di individui dominanti (di cui il 50% di eterozigoti AaBb per entrambi i caratteri ed il 25% di omozigoti dominanti AABB per entrambi i caratteri) ed il 25% recessivi. Non sarebbero ovviamente comparsi i fenotipi con i caratteri scambiati (D/R e R/D) tipici dei caratteri indipendenti.
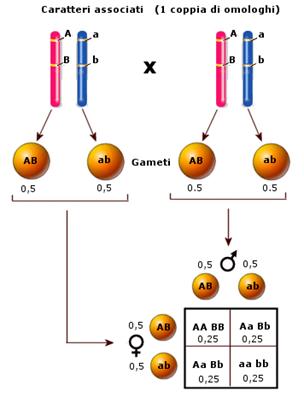
E' possibile verificare se due caratteri sono indipendenti o associati anche reincrociando i diibridi F1 con individui omozigoti recessivi per entrambi i caratteri.
se i caratteri sono indipendenti il diibrido genera 4 tipi di gameti con egual probabilità (1/4) e dal reincrocio si ottengono sia le combinazioni fenotipiche parentali (D/D e R/R) che quelle ricombinanti (D/R e R/D) con egual probabilità (25% ciascuna)

se i caratteri sono associati il diibrido genera solo 2 tipi di gameti con egual probabilità (1/2) e dal reincrocio si ottengono solo le combinazioni fenotipiche parentali (50% D/D e 50% R/R)

Vedremo tuttavia che questa previsione teorica non sempre viene rispettata poiché anche i caratteri associati possono in qualche misura ricombinarsi come conseguenza del processo di crossing-over che avviene durante la meiosi e possono pertanto produrre piccole percentuali di individui ricombinanti.
Si tenga presente che in molti testi di genetica le prime due leggi di Mendel (Legge della dominanza e legge della segregazione) sono riunite in un unico enunciato e che, in questo caso la terza legge di Mendel (legge dell'assortimento indipendente) diventa la seconda.
|
| Appunti su: tabella di mendel, schema del fiore di pisello, 3 legge di mendel tabella, https:wwwappuntimaniacomscientifichechimicamendel-le-leggi-dellereditarie64php, terza legge di mendel immagini, |
|
| Appunti Biologia |  |
| Tesine Geografia |  |
| Lezioni Ingegneria tecnico |  |