|
| Appunti scientifiche |
|
|
| Appunti scientifiche |
|
| Visite: 14356 | Gradito: |
Leggi anche appunti:Numero di ossidazione (nox) o stato di ossidazione (stox)Numero di ossidazione (nox) o stato di ossidazione (stox) Si definisce numero Nomenclatura complessiNomenclatura complessi I metalli di transizione, che allo stato elementare Gli EnzimiGli Enzimi Caratteristiche generali: sono proteine: struttura terziaria |
 |
 |
In alcuni casi un gene può interagire con altri geni per controllare la manifestazione di un unico carattere. L'esperimento classico che ha dimostrato l'esistenza di un'interazione tra geni diversi si deve a Bateson e Punnett (1905) in cui vennero incrociati polli che differivano per la forma della cresta (cresta a rosa x cresta a pisello). In F1 si ottennero un 100% di polli con cresta a noce (o Walnut), un fenotipo nuovo. Poteva trattarsi di un semplice caso di codominanza, con la comparsa di un carattere intermedio. Ma incrociando gli ibridi F1 si ottennero polli con 4 tipi di creste, con una distribuzione di frequenza 9:3:3:1 (9 a noce, 3 a rosa, 3 a pisello e 1 con cresta semplice).
La distribuzione 9:3:3:1 suggeriva che il carattere 'forma della cresta' fosse controllato non da uno, ma due geni secondo il seguente schema
L'allele R da solo determina la cresta a rosa ed è dominante su r
l'allele P da solo determina la cresta a pisello ed è dominante su p
R e P sono codominanti e la loro presenza congiunta determina la comparsa della cresta a noce
rrpp determina la comparsa della cresta semplice.
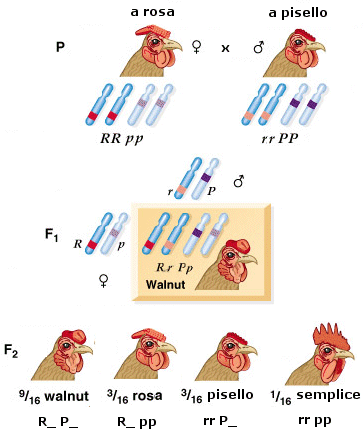
Nella scrittura R_ il trattino significa che il genotipo può essere indifferentemente RR o Rr
Nella scrittura P_ il trattino significa che il genotipo può essere indifferentemente PP o Pp.
Sono stati classificati i seguenti tipi di interazioni tra geni: epistasi, soppressione, complementazione e duplicazione genica.
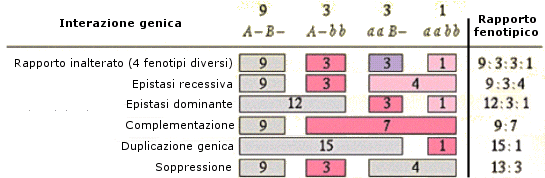
Il risultato di queste interazioni porta in genere ad un cambiamento caratteristico nel rapporto fenotipico (rapporti mendeliani atipici) in F2 rispetto a quello atteso in assenza di interazioni (9:3:3:1).
Si tenga presente che la classificazione delle interazioni geniche viene spesso proposta con criteri diversi, generando non poca confusione.
L'epistasi (leggi epìstasi) è un fenomeno di interazione definito per la prima volta dallo stesso Bateson (1909). Bateson descrisse il fenomeno essenzialmente come una estensione del concetto di dominanza al caso di alleli appartenenti a geni diversi. Il termine viene usato ancor oggi in senso stretto per indicare un effetto di copertura o mascheramento degli effetti di un gene detto ipostatico (letteralmente "che sta sotto") da parte di un altro epistatico (letteralmente "che sta sopra"). Il gene epistatico si manifesta al posto di quello ipostatico. Si tenga tuttavia presente che successivamente il termine epistasi ha allargato il suo significato fino a descrivere in senso lato qualsiasi interazione tra geni diversi che conduca ad una variazione di un unico carattere fenotipico. In molti testi di genetica il termine "interazione epistatica" viene utilizzato come sinonimo di "interazione genica" ad indicare quindi qualsiasi tipo di effetto di un gene su di un altro, contribuendo non poco ad aumentare la confusione su questo argomento.
Noi useremo qui l'accezione più ristretta.
Si riconoscono due tipi di epistasi: dominante e recessiva
Nell'epistasi recessiva l'agente epistatico è un gene in forma omozigote recessiva.
Il rapporto fenotipico caratteristico incrociando i diibridi è 9:3:4
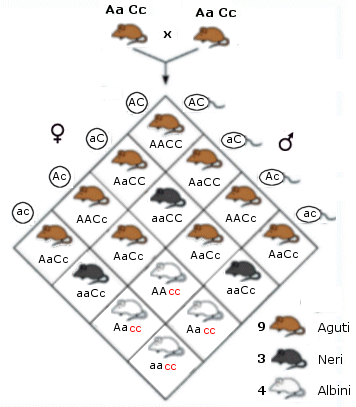
Se ne ha un esempio nel colore del mantello del topo con i due geni aguti ed albino.
Il gene aguti presenta i due alleli
A (dominante) = colore grigio (aguti) a (recessivo) = colore nero
Il gene albino presenta i due alleli
C (dominante) = cromogeno (presenza di colore) c (recessivo) = assenza di colore
Il genotipo omozigote recessivo cc produce albinismo ed è epistatico verso AA, Aa e aa (ipostatici) mascherandone gli effetti. Se incrociamo due diibridi grigi (aguti) AaCc otteniamo infatti
|
Genotipo |
Fenotipo |
Interazione genica |
|
9 A_C_ |
aguti |
Il gene dominante C_ (cromogeno) permette al colore aguti (A_) di manifestarsi |
|
3 A_cc |
bianco |
Il gene epistatico recessivo cc (assenza di colore) si manifesta, mascherando il colore aguti (A_) |
|
3 aa C_ |
nero |
Il gene dominante C_ (cromogeno) permette al colore grigio (aa) di manifestarsi |
|
1 aa cc |
bianco |
Il gene epistatico recessivo cc (assenza di colore) si manifesta, mascherando il colore grigio (aa) |
Nell'epistasi dominante il fattore epistatico è un gene dominante anche in singola dose. Affinché si abbia cioè l'effetto di mascheramento è sufficiente la presenza anche di un solo allele dominante.
Il rapporto fenotipico caratteristico incrociando i diibridi è 12:3:1.
Ne abbiamo un esempio nel colore bianco, nero e marrone del mantello delle pecore.
Anche in questo caso abbiamo due geni, uno che esprime per assenza/presenza di colore e l'altro che esprime per il colore nero/marrone. In questo caso l'allele assenza di colore (B = bianco) è dominante ed epistatico .
B (dominante) = assenza di colore, bianco (epistatico) b (recessivo) = presenza di colore
N (dominante) = colore nero n (recessivo) = colore marrone
L'allele B produce il colore bianco (assenza di colore) ed è epistatico verso NN, Nn e nn (ipostatici) mascherandone gli effetti. Il colore nero (genotipi NN ed Nn) e marrone (genotipo nn) si possono manifestare solo in assenza dell'allele B e quindi quando il gene epistatico è in forma omozigote recessiva (bb), Se incrociamo due diibridi bianchi BbNn otteniamo infatti

|
Genotipo |
Fenotipo |
Interazione genica |
|
9 B_N_ |
bianco |
L'allele B dominante epistatico bianco maschera il colore nero (N_) |
|
3 B_nn |
bianco |
L'allele B dominante epistatico bianco maschera il colore marrone (nn) |
|
3 bb N_ |
nero |
Il gene recessivo bb (cromogeno) permette al nero (N_) di manifestarsi |
|
1 bb nn |
marrone |
Il gene recessivo bb (cromogeno) permette al marrone (nn) di manifestarsi |
Oltre all'epistasi dominante e all'epistasi recessiva, alcuni autori classificano altre tre tipi di epistasi:
doppia dominante
doppia recessiva
doppia dominante-recessiva.
Si tratta di interazioni che possono essere interpretate anche come non epistatiche e di cui parleremo in seguito.
L'interazione di geni complementari (o complementazione) è un fenomeno di interazione fra geni diversi che manifestano la loro azione solo quando sono presenti insieme, generando un fenotipo differente rispetto a quello che generano quando si presentano da soli.
Si riconoscono due tipi di complementazione: dominante e recessiva.
Nell'interazione di geni complementari dominanti (o epistasi doppia recessiva) due geni manifestano un fenotipo diverso quando sono contemporaneamente presenti entrambi almeno con un allele dominante per gene. I rapporti fenotipici caratteristici incrociando i diibridi sono due: 9:7 e 9:6:1.
Un esempio classico di complementazione dominante è quella che abbiamo già analizzato per la forma della cresta dei polli studiata da Bateson e che mantiene il classico rapporto 9:3:3:1, ma con quattro fenotipi differenti.
|
Genotipo |
Fenotipo |
Interazione genica |
|
9 R_P_ |
a noce (Walnut) |
La presenza del gene dominante R_ e del gene dominante P_ genera la cresta a noce |
|
3 R_pp |
a rosa |
La presenza del gene dominante R_ e l'assenza del gene dominante P_ genera la cresta a rosa |
|
3 rr P_ |
a pisello |
L'assenza del gene dominante R_ e la presenza del gene dominante P_ genera la cresta a pisello |
|
1 rr pp |
semplice |
L'assenza del gene dominante R_ e del gene dominante P_ genera la cresta semplice |
A Bateson si devono anche gli incroci su Pisello odoroso (Lathyrus odoratus), che esemplificano il rapporto fenotipico 9:7 .
Nel pisello odoroso la contemporanea presenza dell'allele C e P produce fiori violetti, mentre i genotipi C_pp, ccP_, e ccpp sono bianchi.

|
Genotipo |
Fenotipo |
Interazione genica |
|
9 C_P_ |
Fiori colorati: produzione di antocianina |
Enzimi C e P presenti |
|
3 C_pp |
Fiori bianchi: assenza di antocianina |
Enzima P assente |
|
3 ccP_ |
Fiori bianchi: assenza di antocianina |
Enzima C assente |
|
1 ccpp |
Fiori bianchi: assenza di antocianina |
Enzimi C e P assenti |
La medesima interazione genica legata alla sintesi di antocianina è responsabile anche della colorazione dei semi del mais

Il fenomeno prende anche il nome di epistasi doppia recessiva poiché ogni volta che è presente uno dei due geni in forma omozigote recessiva non è presente il colore. Si potrebbe dunque interpretare il fenomeno come un'azione epistatica del genotipo cc su P_ e del genotipo pp su P_ In realtà si ammette che la formazione del pigmento richieda la presenza contemporanea dell'allele C e P e si spiega attribuendo ad ognuno dei due geni l'azione di sintesi su elementi di una catena enzimatica che partendo da un composto incolore porta ad uno colorato.

L'azione di geni complementari può manifestarsi anche con un rapporto fenotipico 9:6:1.
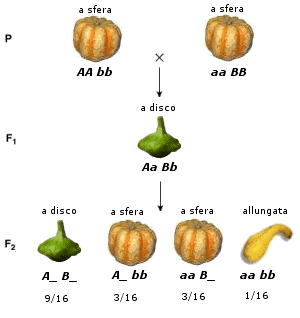
Nella zucchina (Cucurbita pepo), ad esempio, la forma è controllata da due geni.
Il gene dominante A_ ed il gene dominante B_, presenti singolarmente, determinano la forma a sfera.
Il gene dominante A_ ed il gene dominante B_, presenti insieme, determinano la forma a disco. Il gene recessivo aa ed il gene recessivo bb, presenti insieme determinano la forma allungata.
|
Genotipo |
Fenotipo |
Interazione genica |
|
9 A_B_ |
Forma a disco |
La presenza del gene dominante A_ e del gene dominante B_ genera la forma a disco |
|
3 A_bb |
Forma a sfera |
La presenza del gene dominante A_ e l'assenza del gene dominante B_ genera la forma a sfera |
|
3 aa B_ |
Forma a sfera |
L'assenza del gene dominante A_ e la presenza del gene dominante B_ genera la forma a sfera |
|
1 aa bb |
Forma allungata |
L'assenza del gene dominante A_ e del gene dominante B_ genera la forma allungata |
La duplicazione genica (o epistasi doppia dominante o interazione di geni complementari recessivi) è un fenomeno di interazione fra geni diversi che si comportano allo stesso modo, producendo la medesima azione (geni duplicati).
Il rapporto fenotipico caratteristico incrociando i diibridi è 15:1.
Il colore della cariosside del frumento, ad esempio, è controllato da due geni duplicati cromogeni che, in forma dominante, manifestano il medesimo effetto inducendo entrambi la comparsa del pigmento.
Si ipotizza che il pigmento venga sintetizzato da un unico precursore grazie alla presenza di un enzima A o un enzima B o entrambi indifferentemente. La cariosside risulterà pertanto colorata se i due geni A e B che codificano per i rispettivi enzimi, si presenteranno, separatamente o contemporaneamente in forma dominante (A_bb, aaB_, A_B_). La cariosside sarà priva di colore solo se i due geni si presenteranno entrambi in forma recessiva (aabb).
|
Genotipo |
fenotipo |
Attività enzimatica |
|
9 A_B_ |
Cariosside colorata |
Entrambi gli enzimi funzionali |
|
3 A_bb |
Cariosside colorata |
Solo enzima A funzionale |
|
3 aaB_ |
Cariosside colorata |
Solo enzima B funzionale |
|
1 aabb |
Cariosside priva di colore |
Nessun enzima funzionale |

Questo tipo di interazione viene anche interpretata come una interazione di geni complementari recessivi se si considera l'assenza di colore un effetto della compresenza dei due geni allo stato omozigote recessivo (aabb).
Qualche autore la classifica anche come una epistasi doppia dominante poiché il gene dominante A_ può considerarsi epistatico su bb ed il gene dominante B_ può considerarsi epistatico su aa.
La soppressione genica è un fenomeno di interazione fra geni diversi in cui un gene in forma dominante (A_), detto gene soppressore, inattiva un altro gene dominante (B_) permettendo al suo eventuale allele recessivo (nel caso sia presente in forma eterozigote Bb) di esprimersi. Si noti la differenza rispetto ad una epistasi dominante in cui il gene epistatico non permette alla forma recessiva dell'altro gene di esprimersi, ma si esprime al suo posto.
Il rapporto fenotipico caratteristico incrociando i diibridi è 13:3
La sintesi della malvidina nella Primula, ad esempio, è controllata da due geni: un gene dominante K_ che codifica per la sintesi della malvidina ed il gene soppressore dominante D_ che sopprime K e permette, se presente, la manifestazione del suo allele recessivo k (nessuna sintesi di malvidina).
|
Genotipo |
Fenotipo |
Interazione genica |
|
9 K_D_ |
Malvidina non sintetizzata |
L'allele D è presente e l'allele k recessivo può esprimersi |
|
3 K_dd |
Malvidina sintetizzata |
L'allele D è assente e l'allele K dominante può esprimersi |
|
3 kkD_ |
Malvidina non sintetizzata |
L'allele D è presente e l'allele k recessivo può esprimersi |
|
1 kkdd |
Malvidina non sintetizzata |
L'allele D è assente e l'allele K dominante potrebbe esprimersi ma non è presente |
Il rapporto fenotipico 13:3 può essere anche interpretato come una epistasi doppia dominante-recessiva, con il gene dominante A_ del primo locus (epistatico su B_ e bb) che genera lo stesso fenotipo del gene recessivo bb del secondo locus (epistatico su aa) e viceversa
Prendiamo il seguente esempio
Locus 1: A bianco; a colore.
Locus 2: B colore; b bianco.
|
Genotipo |
Fenotipo |
Interazione genica |
|
9 A_B_ |
Bianco |
L'allele A è presente ed essendo epistatico su B_ si manifesta |
|
3 A_bb |
Bianco |
L'allele A è presente ed essendo epistatico su bb si manifesta |
|
3 aaB_ |
Colorato |
L'allele A è assente ed i geni B_ ed aa si manifestano |
|
1 aabb |
Bianco |
il gene bb è epistatico su aa e si manifesta |
|
| Appunti su: interazione genica cresta gallo, interazioni tra geni, interazione doppia, interazione tra geni, https:wwwappuntimaniacomscientifichechimicainterazione-tra-geni85php, |
|
| Appunti Biologia |  |
| Tesine Ingegneria tecnico |  |
| Lezioni Geografia |  |