|
| Appunti universita |
|
|
| Appunti universita |
|
| Visite: 1167 | Gradito: |
Leggi anche appunti:L'Azione Patogena dei BatteriOncogeni ed oncosoppressoriOncogeni ed Oncosoppressori Gli Oncogéni sono geni la cui costante attivazione DermatopolimiositeDERMATOPOLIMIOSITE E un'affezione infiammatorio-degenerativa dei muscoli |
 |
 |
Patterning antero posteriore del sistema nervoso centrale dei vertebrati
A seguito dell induzione neurale si forma dunque già allo stadio di piastra neurale un pattern antero posteriore del SNC rappresentato dall espressione di geni in domini ristretti. Questi geni, alcuni dei quali ho già nominato, sono contemporaneamente causa ed effetto della regionalizzazione del sistema nervoso. La loro presenza in domini specifici è un effetto dell espressione di altri geni che si trovano a monte" nella gerarchia genica dello sviluppo del SNC ad esempio gli inibitori di BMP e i fattori posteriorizzanti , ma è causa della successiva specificazione dei domini in cui sono espressi fino a diventare regioni distinte del SNC definiti i neuromeri ( Shimamura et al. 1995).
Nei vertebrati è possibile ricavare una mappa del destino delle principali regioni del SNC allo stadio di piastra neurale
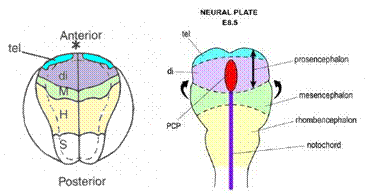 Fig. 1 3 - Mappe del destino
dei territori della piastra neurale di Xenopus e topo
Fig. 1 3 - Mappe del destino
dei territori della piastra neurale di Xenopus e topo
In questo paragrafo descriverò come i geni espressi precocemente a livello di piastra neurale interagiscono per regionalizzare il SN in sviluppo, stabilendo regioni di confine tra i neuromeri e gli organizzatori secondari.
A partire dallo stadio di piastra neurale tardiva, nelle regioni più anteriori si accende il gene Bf1 a delineare le regioni del telencefalo da quelle del diencefalo, prima che si
stabilisca un confine morfologico.
Tanto nei pesci quanto nei mammiferi c è una regione dell ectoderma non neurale, direttamente anteriore alla piastra neurale, che ha la funzione di indurre l espressione di marcatori del telencefalo, le cellule row1" e la anterior neural ridge ANR rispettivamente.
Le cellule della row 1 producono sia Fgf8 che Tlc , un inibitore di Wnt che protegge le regioni più anteriori dall effetto posteriorizzante Otx2 è omologo al gene Orthodenticle Otd ) di Drosophila e codifica per un fattore di trascrizione con omeodominio. E' espresso nel proencefalo e nel mesencefalo dei vertebrati. La delezione di entrambi gli alleli di questo gene nel topo porta alla scomparsa delle strutture anteriori Acampora et al. 199 , che possono essere recuperate se il gene deleto è sostituito con un costrutto in cui la coding region dell ortologo di Drosophila
è inserita tra le sequenze UTR, di Xotx2 che ne regolano la traducibilità del messaggero Acampora et al. 200 .
Il prodotto di Otx2 contribuisce, insieme a Gbx2, a posizionare uno degli organizzatori secondari del SN, posto al confine tra mesencefalo e romboencefalo: l istmo.
Fgf8 oltre ad essere espresso nell ectoderma anteriore al telencefalo è espresso anche nel romboencefalo anteriore e media una parte dell attività induttiva dell istmo: se è espresso ectopicamente nel diencefalo caudale induce l espressione di Fgf8, Wnt1 e En2, creando così un secondo organizzatore dell istmo e facendo acquisire al diencefalo posteriore le caratteristiche di mesencefalo, ma con orientamento opposto rispetto alla normale polarità anteroposteriore ( Crossley et al 19 .
En2 è omologo al gene Engrailed di Drosophila; esso codifica per un fattore di trascrizione con omeodominio. Nei vertebrati ci sono due geni engrailed En1 ed En2
entrambi espressi a cavallo" della regione dell istmo. La differenza tra i due geni è il timing dell espressione, con En1 che è espresso più precocemente rispetto a En2.
Anche i geni Pax2, Pax5 e Pax8 sono necessari per mantenere l identità dell istmo: l istmo è assente in topi knock out per Pax5 e nel mutante noi no isthmus) di zebrafish in cui non è funzionante il gene pax2 1 che è l ortologo dei tre geni pax appena nominati. L espressione di En2 e Wnt1 è attivamente regolata dai geni Pax.
Il gene Gbx2 è espresso nella porzione più anteriore del romboencefalo; il topo Gbx2
- manca del cervelletto, che si origina dalla porzione dorsale dei primi tre rombomeri, e il suo mesencefalo si estende posteriormente Wassarman et al. 199 .
I rombomeri sono gli otto compartimenti r r8) in cui è suddiviso il romboencefalo, l organizzazione segmentale del romboencefalo non si limita alla struttura anatomica esterna, ma anche alla sua organizzazione citologica. Infatti, neuroni che innervano i diversi archi branchiali hanno i loro corpi cellulari all interno di questi rombomeri, distribuiti con una precisa periodicit : in particolare, i corpi cellulari dei neuroni branchiomotori presentano periodicità alternata, quelli degli interneuroni periodicità in registro coi singoli rombomeri.
Esperimenti di trapianti di rombomeri hanno dimostrato che i confini si creano tra rombomeri pari e dispari. Questo accade perché ci sono proprietà adesive diverse tra rombomeri alterni.
I rombomeri non si formano contemporaneamente: prima si formano i confini tra r3/r4 e tra r5 r6, e successivamente i restanti confini.
Sia i geni Hox che geni non Hox come krox20 e Kreisler) sono coinvolti nella suddivisione del romboencefalo. I geni Hox, inoltre, sono coinvolti anche nello stabilire l identità dei vari rombomeri. Krox 20 è espresso già allo stadio di piastra neurale in due aree corrispondenti ai futuri r3 e r5.
In mutanti in cui il gene Krox20 è mutato in entrambi gli alleli mancano r3 e r5 e si forma un territorio in cui i rombomeri pari adiacenti si fondono.
Alla base della formazione dei diversi compartimenti ci sono proprietà adesive e di cell sorting diverse: nei rombomeri si alternano l espressione di efrine nei rombomeri pari), e dei loro recettori nei rombomeri dispari .
L interazione ligando recettore fa iniziare, in entrambe le cellule, una via di segnale che porta alla repulsione delle stesse per cui si creano compartimenti separati.
Krox20 induce l espressione di EphA4 un recettore dell efrina B2.
L identità dei rombomeri è specificata dai geni Hox così come i loro ortologhi nella
Drosophila ne specificano i segmenti.
I geni Hox nei vertebrati contribuiscono anche a creare i segmenti, non solo a attribuirgli un identit .
I geni Hox sono espressi nel SN dei vertebrati , a partire da r2, secondo il principio della colinearità spaziale.
In generale, i domini di espressione dei diversi geni Hox si spostano con una periodicità di due rombomeri: hoxb2 è espresso fino al confine r /r3, Hoxb3 fino al confine r4 r5, Hoxb4 fino al confine r6 r7. Ci sono però delle eccezioni: ad esempio, Hoxb1 è espresso solo in r4.
E' stato osservato nel topo che la soppressione di Hoxb1 fa si che r4 assuma l identità segmentale di r2 cioè le proiezioni dei neuroni brachimotori vanno ad innervare il
primo arco branchiale invece del secondo (Studer et al. 199 ) ; nel pollo invece è
stato fatto l esperimento di guadagno di funzione infettando le cellule di r2 con un retrovirus contenente la sequenza di Hoxb1 in modo che il gene fosse costitutivamente attivo: come conseguenza le cellule di r2 si comportavano come quelle di r4 cioè le proiezioni dei neuroni brachimotori innervano il secondo arco branchiale invece del primo Bell et al. 1 99 . Un guadagno di funzione di un gene Hox porta in genere ad un cambiamento di identità in senso posteriore. I geni Hox sono modulati dall acido retinoico, di cui mediano l effetto posteriorizzant Marshall et al. 1996 .
|
| Appunti su: |
|
| Appunti Nutrizione |  |
| Tesine Bambini |  |
| Lezioni Bellezza |  |